1. Introduction
In the high-stakes arena of global food security, a “Center of Origin” is defined as the geographical area where a group of organisms first developed their distinctive properties. The strategic preservation of plant genetic resources within these Vavilovian centers is not merely a scientific pursuit but a fundamental requirement for maintaining the genetic diversity essential for climate adaptation. For the modern breeder and biodiversity strategist, understanding these origins provides the raw material necessary to bridge the gap between historical evolutionary success and the urgent requirements of contemporary agriculture.
Wild Relatives vs. Domesticated Crops
| Trait Type | Wild/Primitive Form | Domesticated Form |
|---|---|---|
| Seed Retention | Shattering (seeds drop easily to spread) | Non-shattering (seeds stay on plant for harvest) |
| Seed/Fruit Size | Small seeds; often with thick, hard coats | Large seeds or fruits; higher caloric yields |
| Growth Habit | Spreading, trailing, or “weedy” habit | Compact, upright, and uniform habit |
| Adaptation | High seed dormancy; bitter/protective tastes | Reduced dormancy; improved flavor and nutrition |
2. The Vavilovian Foundation: Defining Centers of Origin and Diversity
Nikolai Vavilov revolutionized agricultural strategy by shifting the perspective of crop development from a series of random occurrences to a structured evolutionary process driven by specific geographic foci. He posited that the distribution of genetic wealth is not uniform but clustered in regions where ancient civilizations co-evolved with their staple crops. This framework allows us to distinguish between the “original homes” of crops and the later areas of diversification.
The following table contrasts the two primary classifications identified in the Vavilovian model:
| Feature | Primary Centers of Origin | Secondary Centers of Diversity |
|---|---|---|
| Definition | Original geographic source where a crop evolved from wild progenitors. | Areas where significant diversity developed later through human movement and activity. |
| Allele Frequency | High frequency of dominant alleles (often found toward the center). | High frequency of recessive genes (often found toward the periphery). |
| Morphology | Presence of wild forms and primitive/natural characters. | Prevalence of desirable, domesticated, and improved characters. |
| Selection Type | Primarily driven by natural selection. | Driven by both natural and intensive artificial (human) selection. |
| Genetic Activity | Lower frequency of crossing over. | Higher frequency of crossing over. |
The Strategic Value of Spatial Genetics
Locating dominant genes within these centers is mission-critical for developing resistance to emerging pests and diseases. While the spatial distribution of dominant alleles at the center versus recessive genes at the periphery is a point of ongoing scientific debate, the strategic objective remains clear: these centers harbor the robust genetic blueprints developed during the crop’s initial arms race with its environment. Identifying these dominant traits allows breeders to fortify vulnerable, uniform modern cultivars against catastrophic failure.
As we move from theoretical foundations to the physical map, these centers manifest in specific eco-topographic regions defined by mountainous isolation and tropical or subtropical climates.
3. Global Biogeography: The Eight World Centers of Crop Origin
Vavilov identified that the catalysts for diversification were frequently mountainous regions and tropical/subtropical zones. These diverse eco-topographic conditions provided the isolation and environmental pressure necessary for a vast array of genetic variants to emerge. Through extensive global exploration, Vavilov identified eight primary world centers, which he later refined to include specific sub-centers and endemic counts.
The Vavilovian Centers and Endemic Species
• I. Chinese Center: The largest independent center (138 endemic species). Origin of Soybean, proso millet, opium poppy, and pear.
• II. Indian Center (Hindustan): Divided into two distinct sub-centers:
◦ (a) Indo-Myanmar: (117 species) excluding North-West India. Home to Rice, chickpea, mung bean, and cotton.
◦ (b) Indo-Malayan: (55 species) including the Malayan Archipelago. Home to Banana, coconut, and velvet bean.
• III. Central Asiatic Center: Includes North-West India, Afghanistan, and Uzbekistan (43 species). Key home of Bread wheat, club wheat, pea, and garlic.
• IV. Asia Minor Center: Covering the Near East, Iran, and Turkmenistan (83 species). Origin of Durum wheat, rye, alfalfa, and pomegranate.
• V. Mediterranean Center: Noted for valuable vegetables (84 species). Home to Mediterranean oats, cabbage, and asparagus.
• VI. Abyssinian Center: Centered in Ethiopia and Eritrea (38 species). Source of Sorghum, pearl millet, coffee, and okra.
• VII. Central American Center: Includes South Mexico and Central America (49 species). Origin of Maize, upland cotton, sweet potato, and papaya.
• VIII. South American Center: Includes the Andes, Chile, and Brazil/Paraguay (62 species). Home to Potato, tomato, tobacco, and pineapple. (Sub-centers include Chiloe for potato and Brazil/Paraguay for peanut and rubber).
Vavilov’s map identified eight major geographic regions, often separated by mountains or deserts, where the majority of our food was first domesticated.
| Center | Geographic Region | Representative Crops |
|---|---|---|
| I. Chinese Center | Central & Western China | Soybean, Radish, Peach, China Tea |
| II. Hindustan Center | India, Burma, SE Asia | Rice (Oryza sativa), Sugarcane, Mango, Cotton |
| III. Central Asiatic | NW India, Afghanistan | Bread Wheat (Triticum aestivum), Peas, Garlic |
| IV. Asia Minor | Iran, Turkmenistan | Rye, Pomegranate, Alfalfa, Fig |
| V. Mediterranean | Mediterranean borders | Cabbage, Durum Wheat, Olive, Asparagus |
| VI. Abyssinian | Ethiopia, Somalia | Coffee, Sorghum, Barley, Okra |
| VII. Central American | S. Mexico, Costa Rica | Maize (Zea mays), Guava, Avocado, Papaya |
| VIII. South American | Peru, Chile, Brazil | Potato, Tomato, Tobacco, Pineapple |
Harlan’s “Centers and Non-centers” (1971)
A critical scientific nuance introduced by Jack Harlan is the concept of Non-centers. Unlike monocentric crops with a discernible point of origin, some crops (like Sorghum or Pearl Millet) are noncentric, meaning they were domesticated over a broad geographical span of 5,000 to 10,000 km. This suggests that agricultural development was often a dispersed regional process rather than a single-point event.
The Law of Homologous Series
Vavilov’s Law of Homologous Series suggests that closely related species and genera follow similar patterns of variation. This is most evident in the Fabaceae (Leguminosae) family, where variability in vegetative and seed characters is similar across different genera. This law simplifies the search for new genetic traits; if a variation is discovered in wheat, a similar variation can be predicted and located in its relatives, such as rye or barley.
4. The Functional Reservoir: The Gene Pool Concept
The Gene Pool Concept (Harlan and de Wet) serves as a strategic framework to determine the feasibility of gene transfer. However, for a Senior Strategist, it is vital to acknowledge the technical hurdles: most breeding remains restricted to GP-1 due to undesirable linkage blocks and epistatic interactions found in wild relatives, which can disrupt carefully balanced modern genomes.
The Four-Tier Hierarchy
• GP-1 (Primary): Includes the crop and its interfertile wild/weedy races. Gene transfer is easy, and hybrids are fertile. This is the primary source of variation.
• GP-2 (Secondary): Related species where transfer is possible but difficult. Hybrids are often weak or sterile with poor chromosome pairing.
• GP-3 (Tertiary): Distant relatives where hybridization is extremely difficult, often resulting in lethal or sterile offspring. Requires embryo rescue or bridging species (e.g., using an Agropyron x Triticum derivative to bridge Elymus alleles into wheat).
• GP-4 (Quaternary): Extends the reach of breeding beyond sexual cross-compatibility. This involves synthetic or GMO strains using transformation techniques to move genes across unrelated organisms.
Wide Crosses and Introgressive Hybridization
Strategic “Wide Crosses” allow for introgressive hybridization—the transfer of a small number of genes from one species to another without compromising taxonomic integrity. A premier example is the T1BL.1RS wheat-rye translocation, where a segment of rye chromosome introduced a cluster of genes for resistance to leaf, stem, and stripe rusts, while simultaneously increasing grain yield and kernel weight.
5. Ecosystem Synergy: Biodiversity’s Role in Sustainability
Agriculture is a managed ecosystem dependent on natural biodiversity, which acts as the “pillar of human development.” This synergy is maintained not only by biological processes but also by the tribal wisdom and ethnic knowledge bases of 300–500 million indigenous people.
1. Pollination: Essential for one-third of global crops. The decline of bees, butterflies, and birds directly threatens food security and increases costs via manual pollination.
2. Soil Health: Driven by microbial diversity (bacteria, fungi, algae). These organisms facilitate the cycling of nitrogen, carbon, and phosphorous.
3. Natural Pest Control: Predators provide self-regulating management. Unlike chemical pesticides, which foster resistance and harm non-target microbes, biodiversity provides a stable, healthy ecosystem.
The conservation of these resources is tied to cultural and religious values. In India, the planting and worship of Ocimum sanctum (Tulsi) and Ficus religiosa (Pipal) exemplify how traditional societies preserve biodiversity through belief systems. As noted by Ehrlich and Ehrlich, “humans have an ethical responsibility to protect biodiversity” to maintain this balance.
6. Critical Threats: Genetic Erosion and Economic Risk
Genetic erosion—the loss of traditional landraces and ecotypes—represents a catastrophic risk to future food systems. This loss is not merely environmental; it is a severe economic threat that directly compromises the global trade in herbal formulations and ethno-medicines, which rely on the “raw ingredients of evolution.”
Drivers of Depletion
• Monoculture Practices: The industrial focus on high-performance, uniform cultivars leads to genetic homogeneity, making the entire food system vulnerable to single-point failures.
• GMO Introgression: Unintentional Vertical Gene Transfer from transgenic crops into landraces in centers of origin threatens the genetic integrity of these resources. The Mexican maize case study highlights the risk of “unintentional introgression” into centers of diversity, potentially affecting landrace purity.
• Climate Change: Extreme weather events and shifting temperatures disrupt the synchronization of flowering times and pollinator activity, leading to reproductive “mismatches” in critical crops.
Threats to Global Food Security and the Path Forward
The loss of biodiversity is a direct threat to the “Four Legs of the Chair” that support sustainable development: Economic, Environmental, Social, and Cultural. When we lose biodiversity, the chair collapses. For a farmer, the consequences are immediate:
1. Reduced Yields: Fewer pollinators and poor soil health lead to smaller, lower-quality harvests.
2. Increased Costs: Higher spending on synthetic fertilizers and pesticides to replace lost ecosystem services.
3. Loss of Resilience: Increased vulnerability to “megadisturbances” like climate change and new pests.
4. Nutritional Decline: A lack of genetic variety leads to fewer nutrient-dense food options.
5. Economic Vulnerability: Small-scale farmers face a higher risk of total failure as the natural safety net vanishes.
7. Conclusion: Pathways to Conservation and Resilience
Protecting centers of diversity is a prerequisite for a resilient global food system. The loss of these resources permanently limits our capacity to respond to environmental shifts.
Strategic Conservation Actions
• In situ Conservation: Maintaining landraces on-farm and species in natural habitats (e.g., the Sierra de Manantlan reserve for Zea diploperennis) to allow for continued evolutionary adaptation.
• Ex situ Conservation: Maintaining germplasm in seed banks, botanical gardens, and tissue culture collections as a defensive safety net.
• Integrated Pest Management (IPM): Reducing chemical dependence by leveraging natural predators.
• Agroforestry: Utilizing tree-based systems to provide habitats for pollinators and regulate soil moisture.
Sustainable development is a “chair with four legs”: economic, environmental, social, and cultural. Only by strengthening all four through the conservation of our genetic heritage can we ensure a productive future for global agriculture.
The Three Pillars of Agricultural Insight
• History Matters: Agriculture was a co-evolutionary partnership that transformed human society by allowing for surplus storage and stability.
• Geography Matters: Every crop has a “home address”—a geographic center where its most valuable genetic and cultural secrets are stored.
• Variety is Survival: Genetic diversity is the “raw ingredient” of our survival. It provides the resistance needed to face pests, diseases, and a changing climate.
Image Summary



Questions/Answers
1. How does biodiversity loss impact global food security and agriculture?
Biodiversity loss poses a significant threat to global food security and agricultural sustainability because agriculture is heavily dependent on the diverse natural ecosystems of the planet. The sources identify four critical areas where biodiversity loss negatively impacts agriculture: pollination, soil health, natural pest control, and genetic diversity.
1. Decline in Pollination Services
Pollination is a vital ecosystem service required for the reproduction of many crops, including fruits, vegetables, and nuts.
• Reduced Yields and Quality: Bees alone are responsible for pollinating approximately one-third of the world’s food crops. A decline in pollinator populations—driven by habitat loss, pesticide use, and climate change—leads directly to reduced crop yields and quality.
• Vulnerability of Specific Crops: High-value crops like almonds and blueberries are particularly at risk.
• Economic Impact: The global value of pollination services is estimated to be worth billions of dollars annually; losing these services could lead to increased food prices and the need for expensive alternatives like hand pollination.
2. Degradation of Soil Health
Soil health is maintained by a complex ecosystem of microorganisms, including bacteria and fungi, which support nutrient cycling and water filtration.
• Erosion and Nutrient Loss: Biodiversity loss in soil can lead to soil degradation and erosion, reducing the soil’s capacity to support plant growth. This results in a decrease in crop yields as essential nutrients are lost.
• Reduced Resilience: Biodiverse soil ecosystems have better water-holding capacity and structure. Without these services, agricultural systems become more prone to drought and other climate-related stresses.
• Chemical Dependency: Loss of soil fertility often leads to an increased reliance on synthetic fertilizers, which can cause further environmental damage, such as nutrient pollution in waterways.
3. Loss of Natural Pest Control
Biodiversity provides natural pest control through insects and animals that act as predators to crop-damaging pests.
• Pest Outbreaks: The reduction of natural predators and competitors allows pest populations to grow rapidly, increasing pressure on crops.
• Pesticide Treadmill: As natural pest control declines, farmers often rely more heavily on chemical pesticides. This can lead to a cycle of dependency where pests develop resistance, requiring even higher amounts of chemicals that further harm beneficial insects and soil microbes.
4. Reduction of Genetic Diversity
Genetic diversity is the “raw ingredient” for crop improvement and evolution.
• Loss of Adaptation: Genetic diversity allows for the breeding of new crop varieties that are resistant to diseases and adapted to changing environmental conditions, such as climate change.
• Genetic Vulnerability: Modern agriculture’s focus on a limited number of high-yielding varieties (monocultures) creates genetic homogeneity, making the entire food system vulnerable to catastrophic failure if a new disease or pest emerges. For example, the Irish potato famine was a result of a narrow genetic base.
• Nutritional Impact: A lack of genetic diversity in crops can lead to reduced nutritional diversity in human diets, impacting global health.
Broader Socio-Economic Consequences
The cumulative effect of biodiversity loss in agriculture extends to human well-being and economic stability:
• Food Insecurity: In developing countries, where many rely on subsistence agriculture, the decline in ecosystem services like pollination and soil fertility can lead directly to increased poverty and malnutrition.
• Loss of Traditional Knowledge: The disappearance of traditional crop and livestock varieties also means the loss of unique cultural and historical knowledge associated with them.
• Reduced Resilience: Overall, the loss of biodiversity reduces the ability of the global food system to adapt to extreme weather events and new biological threats.
To mitigate these impacts, the sources emphasize the importance of adopting agroecological practices, such as crop rotation, integrated pest management, and the conservation of traditional varieties, to build a more resilient and sustainable food system.
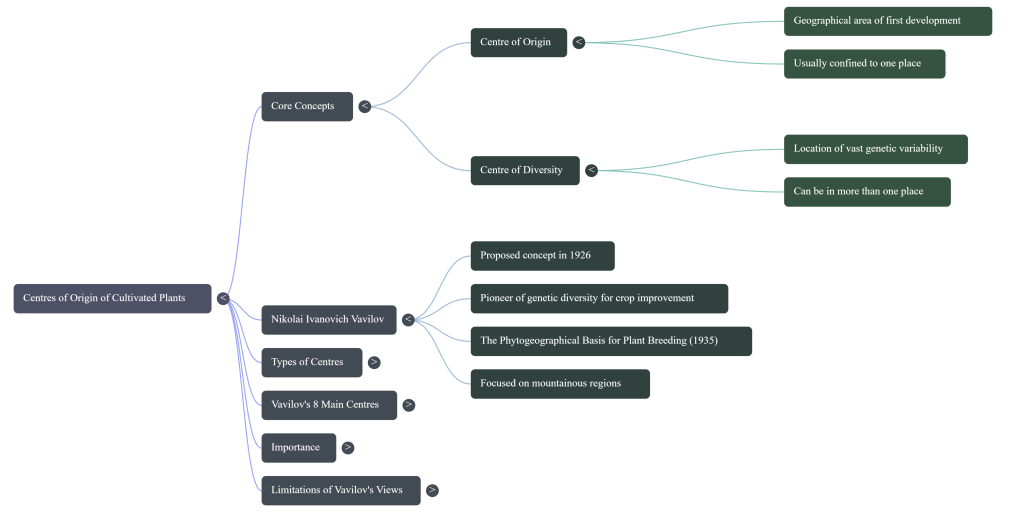
2. What is Vavilov’s theory of crop origins and its limitations?
Nikolai Vavilov’s theory of crop origins, first proposed in his 1926 work Studies on the Origin of Cultivated Plants, posits that each crop has a characteristic primary center of diversity that is also its center of origin. Vavilov believed that these centers were localized areas—often isolated by mountains, deserts, or seas—where genetic variation accumulated over time through mutations and subsequent selection.
Core Concepts of Vavilov’s Theory
Vavilov distinguished between different types of diversity centers:
• Primary Centers of Origin: These are the original homes of a crop where it evolved from wild species. They are characterized by vast genetic diversity, a high frequency of dominant alleles, and the presence of wild relatives.
• Secondary Centers of Diversity: These are regions where a crop shows considerable diversity but did not originally evolve. These areas are often associated with human activities and are characterized by recessive alleles and traits desirable for cultivation.
• Microcenters: Small areas within larger centers of diversity that exhibit an exceptionally high rate of natural evolution and genetic richness.
• Law of Homologous Series: Vavilov also proposed that closely related species and genera tend to exhibit similar series of genetic variation, which simplifies the study of related plants.
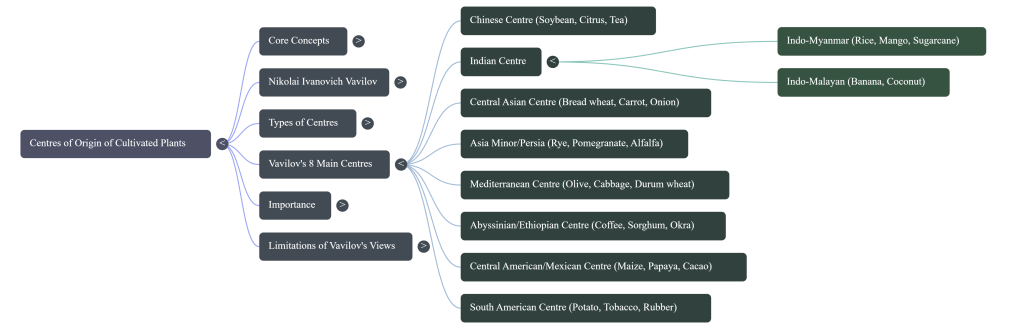
The Eight World Centers of Origin
Vavilov eventually identified eight main geographic regions where major crops were domesticated:
1. Chinese Center: The largest independent center; origin of soybean, radish, pear, and peach.
2. Hindustan Center (India/Southeast Asia): Origin of rice, sugarcane, mung bean, and citrus.
3. Central Asiatic Center: Origin of common wheat, peas, lentils, and garlic.
4. Asia Minor Center (Near East): Origin of several wheat species, rye, pomegranate, and fig.
5. Mediterranean Center: Origin of olives, cabbage, durum wheat, and many vegetables.
6. Abyssinian Center (Ethiopia): Origin of coffee, sorghum, pearl millet, and okra.
7. Central American Center: Origin of maize, common bean, cotton, and papaya.
8. South American Center: Origin of potatoes, tomatoes, tobacco, and pineapple.
Limitations of Vavilov’s Theory
Modern scientific expansion has identified several limitations to Vavilov’s original views:
• Diversity Does Not Always Equal Origin: It is now known that the region with the greatest genetic diversity is not always the center of origin. For instance, modern studies suggest that gene flow from ancestors can shift the center of diversity far from the actual site of domestication, as seen in emmer and common wheat.
• Geographic Scope: Vavilov believed domestication occurred almost exclusively in tropical and subtropical mountainous regions, but evidence now suggests many crops originated in plains.
• Allele Frequency Patterns: His assertion that dominant alleles are always concentrated at the center while recessive alleles move toward the periphery is no longer accepted as a universal rule.
• Multiple Origins: Recent evidence indicates that many crops have multiple centers of origin rather than a single point of domestication.
• Omitted Regions: Vavilov did not adequately cover Africa or Australia, both of which possess significant wealth in crop genetic diversity.
• Lack of Evidence: For many species, the exact origin cannot be traced due to insufficient archaeological or historical evidence.
3. How do gene banks and ex situ conservation protect biodiversity?
Gene banks and ex situ conservation protect biodiversity by preserving genetic resources outside of their natural habitats, ensuring that the genetic wealth of species remains available for future generations, research, and agricultural improvement. These methods act as a safeguard against genetic erosion—the loss of diversity due to habitat destruction, urbanization, and the replacement of traditional landraces with modern, uniform cultivars.
Mechanisms of Ex Situ Conservation
Ex situ conservation involves removing organisms or genetic material from their original environment to be maintained in specialized facilities. The sources describe several key methods:
• Seed Banks: This is the most common and cost-effective method for seed-propagated species. Seeds are dried to specific moisture levels and stored in moisture-proof containers at subfreezing temperatures, which can preserve their viability for 50 years or more.
• Field Gene Banks and Botanic Gardens: For plants that are vegetatively propagated or produce “recalcitrant” seeds (seeds that cannot survive drying), full plants are grown in field banks or arboreta. Over 12,000 threatened plant species are currently maintained in more than 1,500 botanical gardens worldwide.
• In Vitro and Cryopreservation: Clonal species and those with poor seed production are maintained using tissue culture (suspension cells, callus, or meristems). Cryopreservation involves storing cells or vegetative material in liquid nitrogen at extremely low temperatures (-150°C to -196°C) to maintain genetic integrity indefinitely.
• Culture Collections and Aquaria: Laboratories maintain collections of microorganisms (bacteria, fungi, viruses, and algae), while aquaria are utilized to conserve aquatic organisms.
Role of Gene Banks in Protecting Biodiversity
Gene banks serve as genetic repositories that store a wide range of materials, including landraces, obsolete cultivars, advanced breeding lines, and wild relatives,,. They protect biodiversity through the following functions:
• Mitigating Genetic Vulnerability: By maintaining diverse genetic stocks, gene banks provide the “raw ingredients” needed to breed new varieties resistant to pests, diseases, and environmental stresses,. This helps prevent “genetic disasters,” such as the Irish Potato Famine, which occurred because of a narrow genetic base,.
• Supporting Agricultural Sustainability: Biodiversity is essential for food security,. Gene banks allow breeders to access specific genes—often from wild relatives—to improve crop yields and adapt agriculture to changing climates,,.
• Preserving “Insurance” Samples: Gene banks often divide their holdings into base collections (for long-term “insurance” storage), active collections (for distribution to researchers), and working collections (for immediate breeding use),.
Strategic Importance
Because it is impossible to conserve all plant genetic diversity solely in their natural habitats (in situ), ex situ conservation provides a necessary, static, and often safer backup,. Large-scale international efforts, such as the 15 gene banks managed by the Consultative Group on International Agricultural Research (CGIAR), maintain hundreds of thousands of samples of essential staple crops to ensure they remain a public good for all humanity,. Additionally, the Svalbard Global Seed Vault serves as a global “base” gene bank to protect these reserves against large-scale regional or global crises.
4. What is the Irish potato famine example?
The Irish potato famine of the 1840s is cited in the sources as one of the most devastating historical examples of the dangers of genetic vulnerability and the importance of maintaining a broad genetic base in agriculture.
The famine resulted from the following factors:
• Reliance on a Single Variety: During this period, the Irish population was heavily dependent on a single potato variety known as the Lumper.
• Genetic Homogeneity: Because potatoes are vegetatively propagated, all Lumper potatoes were genetically identical. This created a narrow genetic base, meaning the entire national crop had the same strengths and, critically, the same weaknesses.
• The Pathogen: The famine was triggered by the introduction of the late blight pathogen (Phytophthora infestans, specifically mating type A1) from another geographical region. Because the Lumper variety lacked any natural resistance to this specific pathogen, the disease was able to spread rapidly and uncontrollably.
• Human and Economic Impact: The total destruction of the potato crop led to catastrophic food shortages, resulting in the starvation and death of more than one million people in Ireland.
This example is frequently used to illustrate the risks of monoculture and the loss of biodiversity. It highlights why modern agricultural strategies prioritize the conservation of landraces and wild relatives, as these diverse genetic resources provide the necessary “raw ingredients” to breed resistance into modern crops and prevent similar catastrophic failures in the future.
5. Explain agroecological practices like crop rotation.
Agroecology is a science-based approach that seeks to enhance the natural ecological processes that support agriculture while simultaneously minimizing negative environmental impacts. Agroecological practices are designed to integrate biodiversity into agricultural ecosystems to build a food system that is more sustainable, resilient, and productive.
The sources highlight several key practices used to achieve these goals:
Crop Rotation
Crop rotation is a central cultural practice that involves alternating the types of crops grown in a specific area across different seasons.
• Soil Health and Fertility: One of its primary functions is to maintain soil health and fertility by promoting nutrient cycling and preventing the depletion of specific soil elements that occurs when the same crop is grown repeatedly.
• Soil Structure: By rotating crops, farmers can increase soil organic matter and enhance soil structure, leading to better water infiltration and reduced compaction.
• Natural Pest Control: Rotation serves as an effective natural pest management tool because it disrupts the life cycles of specific pests and diseases that might otherwise thrive in a monoculture environment.
• Erosion Prevention: It helps prevent soil erosion by ensuring the ground is managed in a way that promotes aggregation and porosity.
Other Related Agroecological Practices
Agroecology often employs a combination of strategies to support biodiversity:
• Cover Cropping: Often used alongside rotation, cover crops are planted primarily to protect and enrich the soil between main growing seasons rather than for harvest. They prevent erosion, promote nutrient cycling, and provide habitats for beneficial insects.
• Intercropping: This involves growing two or more crops simultaneously in the same field, such as legumes and grains. This practice provides diverse habitats for natural predators, such as spiders and ground beetles, which help control pest populations naturally.
• Agroforestry: This approach integrates trees and perennial crops into agricultural landscapes. Trees help regulate moisture, provide wildlife corridors, and promote the storage of soil carbon.
• Conservation Tillage: By practicing minimal soil disturbance, farmers protect the diverse communities of microorganisms, fungi, and bacteria that are essential for nutrient cycling and healthy soil biota.
• Integrated Pest Management (IPM): IPM is a comprehensive strategy that prioritizes cultural and biological methods—such as crop rotation and the promotion of natural predators—over chemical ones. It aims to manage pests with a focus on minimizing harm to non-target species like bees and butterflies.
By adopting these practices, farmers can reduce their dependency on synthetic fertilizers and pesticides, which often lead to nutrient pollution in waterways and the development of pesticide-resistant pests. Ultimately, the sources suggest that these agroecological methods are essential for mitigating the negative impacts of biodiversity loss and ensuring global food security.
6. How do pollinators reduce pest vulnerability?
According to the sources, pollinators reduce pest vulnerability primarily by maintaining the health of crops and ecosystems and facilitating genetic diversity. While pollinators like bees, butterflies, and birds are most famous for their role in crop reproduction, their contribution to pest management is multi-faceted:
1. Maintaining Genetic Diversity
Pollination is a critical mechanism for maintaining genetic diversity in both crops and wild plants. This genetic variation is the “raw ingredient” that allows species to adapt to changing environments and evolve natural resistance to pests and diseases.
• Reduced Vulnerability: When pollinators facilitate cross-pollination, they help prevent genetic homogeneity (monocultures). A diverse genetic base ensures that if a new pest emerges, the entire crop population is less likely to be wiped out, as some individuals may possess natural defenses.
• Consequences of Loss: The sources note that a decline in pollinator populations leads directly to reduced genetic diversity, which increases the vulnerability of agricultural systems to catastrophic pest infestations.
2. Ecosystem Resilience and Health
Pollinators are vital components of a balanced ecosystem. Their presence supports the overall resilience of the agricultural landscape.
• Natural Balance: Biodiverse systems are more resilient to environmental stresses and biological threats than simplified monocultures. By supporting the reproduction of various plant species, pollinators ensure a stable environment where pest populations are less likely to experience unchecked growth.
• Stress Reduction: Healthy, well-pollinated crops are generally more robust and better able to withstand the pressure of pest damage.
3. Synergistic Habitat Benefits
The sources highlight that the habitats required for pollinators—such as flowering plants, meadows, and hedgerows—also support other beneficial insects that provide natural pest control.
• Supporting Natural Predators: Flowering plants provide food and shelter for beneficial insects like ladybugs, lacewings, and parasitic wasps. These insects act as natural predators, preying on crop-damaging pests and reducing the need for chemical pesticides.
• Integrated Pest Management (IPM): Agroecological practices often involve planting “insectary strips” or wildflowers to attract these beneficial organisms. By fostering a landscape that supports pollinators, farmers simultaneously promote a community of natural enemies that keep pest populations in check.
In summary, pollinators serve as a defensive layer for agriculture by ensuring crops remain genetically diverse and physically robust, while the habitats that sustain them also harbor the natural predators necessary for biological pest control.
7. What are the key differences between in situ and ex situ conservation?
The primary difference between in situ and ex situ conservation lies in whether genetic resources are preserved within or outside of their natural environment. While in situ conservation focuses on protecting species in their original habitats to allow for continued evolution, ex situ conservation provides a controlled, static backup to prevent the immediate loss of genetic diversity.
The key differences identified in the sources are summarized below:
1. Location and Setting
• In situ (On-site): This approach maintains genetic resources within the evolutionary dynamic ecosystems of their original habitat. It includes natural parks, biosphere reserves, and “gene sanctuaries”. It also encompasses on-farm conservation, where farmers maintain landraces in the fields where they developed their distinctive properties.
• Ex situ (Off-site): This involves removing genetic resources (seeds, organisms, or individuals) from their original habitat to be maintained in specialized facilities. Examples include seed banks, botanical gardens, arboreta, and laboratories for tissue culture or cryopreservation.
2. Evolutionary Dynamics
• In situ is dynamic: It allows for ongoing adaptation and evolutionary processes to continue in response to changing environmental conditions, pests, and diseases.
• Ex situ is static: It is a static approach designed to “freeze” the genetic makeup of a sample at the time of collection. While this prevents genetic change, it also means the stored material does not adapt naturally to new environmental pressures.
3. Management and Cost
• In situ challenges: Managing natural reserves can be costly and problematic. These areas are often difficult to maintain and can only cover a small portion of a species’ total distribution.
• Ex situ efficiency: Seed-based ex situ conservation is considered a cheap and effective method. It is easy to maintain, requires minimal space, and allows for easier handling and distribution of germplasm for research and breeding.
4. Security and Scope
• In situ risks: Resources in the wild are subject to environmental stresses, habitat destruction, and illegal harvesting.
• Ex situ safety: It acts as a safer backup against the total extinction of a species in the wild. However, it is limited by the physical capacity of gene banks and the difficulty of storing “recalcitrant” seeds that cannot survive traditional drying and freezing.
Summary Table of Differences
| Feature | In Situ Conservation | Ex Situ Conservation |
|---|---|---|
| Location | On-site (natural habitat or farm) | Off-site (gene bank, garden, lab) |
| Nature | Dynamic; allows for continued evolution | Static; preserves genetic integrity |
| Examples | National parks, gene sanctuaries, on-farm | Seed banks, cryopreservation, botanical gardens |
| Cost | Generally higher/more complex | Generally lower for seed storage |
| Main Goal | Maintaining ecosystem processes | Safeguarding germplasm for future use |
These methods are complementary; effective biodiversity protection requires combining the dynamic, evolutionary benefits of in situ conservation with the high security of ex situ gene banks.
8. How do gene banks use cryopreservation for long-term storage?
Gene banks use cryopreservation (also known as freeze-preservation) as a highly specialized method for the long-term conservation of “base collections,” which are intended to be stored as insurance for 50 years or more.
This process involves the following key mechanisms and applications:
Mechanism and Temperature
Cryopreservation involves storing biological materials at extremely low temperatures, typically ranging from -150°C to -196°C. The material is placed in liquid nitrogen, which effectively “freezes” biological activity and prevents genetic change over time.
Materials Targetted for Cryopreservation
While many crops can be stored as dried seeds in traditional seed banks, cryopreservation is essential for specific types of genetic resources that cannot be preserved through standard methods:
• Vegetatively Propagated (Clonal) Species: This includes crops like potatoes, yams, cassava, and sweet potatoes, which have poor seed production or are traditionally grown from vegetative parts.
• Recalcitrant Seeds: Some species produce seeds that lose viability if their moisture content drops below 12% to 30% (e.g., coconut, coffee, cocoa, and rubber). These species are instead maintained using in vitro techniques like tissue culture or cryopreservation of embryos.
• Specific Plant Tissues: Gene banks use cryopreservation for plant cells, tissues, suspension cells, callus, and meristematic tissues.
The Use of Meristem Cultures
Meristem cultures are favored for cryopreservation for several reasons:
• Pathogen-Free Storage: They allow genotypes to be conserved indefinitely while remaining free from pathogens.
• Genetic Integrity: Meristems maintain the genetic integrity of the plant, ensuring that the regenerated plant is an exact match to the original.
• Speed of Regeneration: While forest or fruit trees can take 10 to 20 years to reach maturity via seed, meristems can be subcultured and regenerated into full plants much more rapidly.
Strategic Role in Conservation
In the global network of gene banks, cryopreservation serves as a static form of ex situ conservation. It is often used for “base collections,” which are only accessed for regeneration purposes when germplasm from active or working collections is unavailable. This ensures that even if a species becomes threatened in its natural habitat (in situ), a secure, disease-free genetic backup remains available for future breeding and research.
9. What role does the Svalbard Global Seed Vault play in global security?
The Svalbard Global Seed Vault serves as the world’s global “base” genebank, playing a critical role in global food security by providing a ultimate safety net for agricultural biodiversity. Located in Norway, it is designed to provide long-term secure storage for duplicates of seed samples held in other genebanks across the planet.
Its specific roles in safeguarding global security include:
• Protecting Global Food Reserves: The vault’s primary mission is to protect the food reserves for all humanity by ensuring that the genetic “raw material” of our food supply is never lost.
• Acting as a Secure Back-up: It houses more than 774,000 accessions donated from 53 international genebanks, representing 156 distinct crop genera. This ensures that if a regional genebank’s collection is destroyed by disaster, war, or equipment failure, the genetic resources can be recovered.
• Preserving Genetic Diversity for the Future: By storing a vast array of species—including essential staple crops, legumes, and cereals—it ensures that the genetic wealth required to breed new, resilient crop varieties remains available for future generations.
• Ensuring Public Access: The accessions within these collections are considered an international public good, maintained under the terms of the International Treaty on Plant Genetic Resources for Food and Agriculture (ITPGRFA) to support global research and breeding efforts.
In summary, the vault acts as a Genetics of Disaster safeguard, mitigating the risks of genetic erosion and ensuring that the foundation of our agricultural system is resilient against global crises.
10. How does gene flow shift a crop’s diversity center?
According to the sources, gene flow—specifically the transfer of genetic material from an ancestor to a crop after its initial domestication—is the primary cause for the shift of a crop’s diversity center away from its center of origin. This process creates a “diversity paradox” where a crop may exhibit higher genetic variation in a region far from where it was first domesticated.
The Mechanism of Displacement
The sources explain that shifts in diversity centers typically occur under the following conditions:
• Peripheral Domestication: If a crop is domesticated in a peripheral or specialized population of its wild ancestor, it initially starts with a limited genetic base.
• Sympatry and Introgression: As the crop spreads geographically, it may become sympatric (living in the same area) with more diverse populations of its wild or domesticated ancestors.
• Altering Diversity Patterns: Continuous gene flow (introgression) from these diverse ancestral populations into the crop increases the crop’s genetic variation in that new region. Consequently, the new region becomes a center of diversity, while the actual center of origin remains a region of lower diversity.
Key Examples of Diversity Shifts
The sources provide two detailed examples in wheat species where gene flow has shifted diversity centers:
• Tetraploid Emmer Wheat:
◦ Center of Origin: Molecular studies suggest emmer was domesticated in the Diyarbakir region of southeastern Turkey.
◦ Center of Diversity: Its current highest diversity is found in the Mediterranean basin.
◦ Reason for Shift: Domesticated emmer in the Mediterranean was sympatric with the most diverse wild emmer populations. Gene flow from these wild populations increased the diversity of the domesticated varieties in that region, causing the diversity center to overlap with the wild progenitor’s diversity center rather than the crop’s origin site.
• Hexaploid Common Wheat (Triticum aestivum):
◦ Center of Origin: Its putative origin is in Transcaucasia and northwestern Iran.
◦ Center of Diversity: Its center of molecular diversity is currently in Turkey.
◦ Reason for Shift: Common wheat underwent prolonged sympatry with domesticated and wild tetraploid wheats in Turkey. Continuous gene flow via hybridization (often through pentaploid hybrids) from these tetraploid ancestors served as the principal source of new genetic diversity for the common wheat’s A and B genomes in that region.
Scientific Implications
These findings challenge Nikolai Vavilov’s original hypothesis that the center of highest diversity is always the center of origin. Modern molecular studies indicate that the geographical center of crop diversity may only coincide with the center of origin if domestication occurred directly within the ancestor’s own center of diversity. Otherwise, subsequent gene flow is likely to displace the maximum diversity relative to the site of origin.
11. What are the risks of using GMOs in centers of diversity?
Using genetically modified organisms (GMOs) in centers of diversity—the geographic regions characterized by high levels of genetic variation for specific crops and their wild relatives—poses several significant risks to agricultural biodiversity and conservation efforts.
According to the sources, the primary risks include:
1. Unintentional Introgression and Gene Flow
The most frequently cited concern is the potential for unintentional introgression, where transgenes from GM crops move into traditional landraces or wild and weedy relatives through cross-pollination.
• Reproductive Biology: The risk is highest for out-crossing species like maize and lower for self-pollinated crops like soybean or rice.
• High-Risk Crops: Some crops, such as sorghum, are classified as high-risk because they hybridize easily with pervasive weeds like Johnson grass.
• Genetic Stability: While hybridization is a one-time event, introgression requires repeated backcrossing and the stabilization of the transgene in the host genome, which can permanently alter the genetic makeup of native populations.
2. Ecological Impacts and Increased Fitness
If a transgene confers a survival advantage (such as resistance to pests, diseases, or environmental stress), it could enhance the ecological fitness of wild or weedy relatives.
• Invasiveness: Recipient plants could establish populations that out-compete other native species in the same community, potentially leading to increased weediness or the displacement of wild relatives.
• Trophic Cascades: Such imbalances can lead to unforeseen changes in the local ecosystem, affecting natural biological balances.
3. Genetic Erosion and Replacement of Landraces
Similar to the “Green Revolution” impact, the introduction of GM varieties can lead to the replacement of traditional landraces.
• Market Pressure: Farmers may abandon diverse local varieties in favor of high-performing GM varieties to meet market trends, causing the loss of unique genetic traits adapted to local environments.
• Loss of Integrity: In centers like Mexico (for maize) or the Andes (for potatoes), the introduction of GM varieties threatens the integrity of traditional agricultural systems that have evolved over millennia.
4. Contamination of Germplasm Collections
GMOs pose a direct threat to the management of gene banks and germplasm collections.
• Collection Risks: Collectors may unknowingly gather samples already exposed to GM pollen or mechanical mixtures in markets or farmers’ fields.
• Regeneration Risks: During the mandatory regeneration of seed samples in the field, accessions may be exposed to gene flow if GM crops are grown in nearby fields.
• Uncertainty for Researchers: The unintentional presence of transgenes in germplasm collections can complicate scientific research and the international exchange of genetic resources.
5. Legal and Intellectual Property Issues
The presence of patented GM traits in traditional landraces or gene bank accessions creates complex Intellectual Property Rights (IPR) and legal challenges.
• Ownership Conflicts: Transgene introgression can lead to legal disputes over the ownership of “contaminated” seeds, as seen in various lawsuits involving patent infringement.
• Restrictive Access: IPR protections can limit the ability of small-scale farmers and researchers to access and share genetic resources, potentially hindering future crop improvement efforts.
To mitigate these risks, the sources recommend rigorous biosafety measures, including strict isolation distances during regeneration, routine testing of incoming germplasm, and regional collaboration to monitor the movement of GM varieties within centers of diversity.
12. Explain the role of the Svalbard Global Seed Vault.
Based on the sources, the Svalbard Global Seed Vault in Norway serves as the world’s global “base” genebank, acting as a final safety net for agricultural biodiversity.
Its primary roles in global security and conservation include:
• Ultimate Backup for Global Genebanks: The vault provides long-term secure storage for duplicates of seed samples held in other collections around the world. It currently houses more than 774,000 accessions donated from 53 international genebanks, representing 156 distinct crop genera.
• Protection of Food Reserves: Its fundamental mission is to protect the food reserves for all humanity. By safeguarding these genetic resources, it ensures that the “raw material” needed for crop improvement and adaptation remains available even if local or regional collections are destroyed.
• Safeguard Against “Genetics of Disaster”: The vault is a critical tool for mitigating genetic erosion—the loss of wild diversity and localized landraces—and protecting against “genetic wipeout” caused by natural disasters, war, or large-scale environmental changes.
• Preservation of Invaluable Traits: It preserves seeds with specific desired traits to prevent their extinction, ensuring that genetic variability—the basis of all plant breeding—is maintained for future humanitarian use.
In the broader context of germplasm conservation, the Svalbard facility functions as a base collection, which the sources define as a collection intended for long-term preservation (50 years or more). These collections are typically stored at subfreezing temperatures and are primarily accessed only for regeneration when germplasm from active or working collections is no longer available.
13. How do gene flow and modern genetics challenge Vavilov’s theories?
Modern genetics and the study of gene flow have significantly challenged Nikolai Vavilov’s original theories by demonstrating that the geographical center of a crop’s diversity does not necessarily coincide with its center of origin. While Vavilov’s pioneering work established the importance of genetic diversity, modern research reveals that the evolutionary history of crops is far more complex than a single point of origin radiating outward.
The Role of Gene Flow in Shifting Diversity Centers
The sources identify gene flow as the primary cause for the discrepancy between where a crop originated and where it is currently most diverse.
• Displacement of Diversity: If a crop is domesticated from a peripheral or specialized population of its wild ancestor, its initial genetic base is narrow. However, as the crop spreads and becomes sympatric (occupying the same area) with more diverse ancestral populations, continuous gene flow (introgression) from those ancestors can dramatically increase the crop’s diversity in that new region.
• The Case of Emmer Wheat: Molecular studies suggest emmer wheat was domesticated in the Diyarbakir region of southeastern Turkey. However, its current center of diversity is the Mediterranean basin. This “diversity paradox” occurred because domesticated emmer in the Mediterranean was sympatric with highly diverse wild emmer populations, and gene flow from these wild relatives shifted the crop’s diversity center away from its Turkish site of origin.
• Hexaploid Common Wheat: Similarly, common wheat likely originated in Transcaucasia and northwestern Iran, but its center of molecular diversity is in Turkey. This is due to prolonged sympatry and continuous gene flow from domesticated and wild tetraploid wheats in Turkey.
Challenges from Modern Genetic Evidence
Modern genetic tools, such as molecular markers, RFLP genotyping, and DNA sequencing, have overturned several of Vavilov’s fundamental assumptions:
• Allele Frequency Patterns: Vavilov proposed that primary centers are marked by a high frequency of dominant alleles, while recessive alleles increase toward the periphery. Modern genetics has found this view to be unacceptable and scientifically unsupported as a universal rule.
• Multiple Origins: Rather than a single “center,” modern evidence indicates that many crops have multiple independent centers of origin. For example, common beans (Phaseolus vulgaris) show evidence of multiple domestication events in both Central America and the Andean region.
• Diversity without Origin: In some cases, crops like maize and tomato exhibit vast genetic diversity in regions far removed from their actual centers of origin.
Theoretical Limitations Identified by Modern Research
14. Beyond gene flow, the sources note other areas where Vavilov’s theories have been modified:
• Geographic Scope: Vavilov believed domestication occurred almost exclusively in tropical and subtropical mountainous regions. Modern archaeological and genetic evidence suggests that plains were also significant centers of origin for many cultivated plants.
• Omitted Regions: Vavilov’s original work did not adequately cover the vast genetic wealth found in Africa and Australia, both of which are now recognized as critical areas for crop diversity.
• “Non-centers”: Some scientists, like Jack Harlan, proposed that agriculture did not always develop in a defined “center” but rather in broad “non-centers” where domestication activities were dispersed over thousands of kilometers.
In summary, while Vavilov’s work remains a “founding pillar” for understanding plant genetic resources, modern genetics distinguishes between a center of origin (where a species first appeared) and a center of diversity (where it exhibits the highest variation), acknowledging that environmental factors and human-mediated gene flow often separate the two.
15. What is the Law of Homologous Series?
The Law of Homologous Series (also referred to as the law of homologous variation) was proposed by the Russian scientist Nikolai Vavilov in 1920. The core principle of this law is that closely related species and genera are characterized by similar series of inherited variation.
Key aspects of this theory include:
• Predictive Value: Because related species tend to vary in similar ways, the law makes the study and collection of genetic resources simpler. By observing the variations in one species, a researcher can predict that similar variations or traits likely exist in a closely related species or genus.
• Examples in Nature:
◦ Fabaceae (Leguminosae) Family: Vavilov observed that different genera within this family exhibited similar variations in their vegetative organs, flowers, fruits, and seeds.
◦ Wheat: Identical series of characters are found across different ploidy levels, including diploid, tetraploid, and hexaploid wheat.
◦ Cotton: Similar patterns of variation are noted between diploid and tetraploid cotton species.
• Modern Significance: Vavilov’s concept remains a fundamental principle in agricultural science. Today, it is applied to the field of molecular genetics and is considered a “dogma” in plant breeding and evaluation.
In the broader context of Vavilov’s work, this law was developed alongside his theories on centers of origin and genetic erosion to help systematize the search for valuable plant traits for crop improvement.
16. What is the primary difference between primary and secondary centers?
The primary difference between primary and secondary centers lies in whether the region is the original site of a crop’s evolution from wild species or a later region where diversity developed due to human activity and migration.
According to Vavilov’s theories and the provided sources, the distinctions can be categorized as follows:
1. Origin and Development
• Primary Centers of Origin: These are the original homes of a crop where it first evolved from its wild ancestors. They are typically located in uncultivated areas such as mountains, hills, river valleys, and forests.
• Secondary Centers of Diversity: These are geographic areas where a crop exhibits considerable diversity but did not originally evolve. These centers developed primarily due to human activities, such as migration and the crossing of two or more species.
2. Genetic Characteristics
• Primary Centers: These regions are characterized by a high frequency of dominant alleles and “wild” characters. They typically exhibit less genetic “crossing over” because natural selection is the dominant force.
• Secondary Centers: These regions often contain a large number of recessive genes and traits that are “desirable” for human use and cultivation. Because these areas are generally cultivated, they exhibit more crossing over and are shaped by both natural and artificial selection.
3. The “Diversity Paradox” and Modern Shifts
While Vavilov originally believed that a crop’s primary center was always its center of highest diversity, modern research has identified a “diversity paradox” where secondary centers can become more diverse than primary ones.
• Gene Flow: Diversity can increase in secondary centers away from the point of origin due to gene flow (introgression) from wild or domesticated ancestors that the crop encounters as it spreads into new regions.
• Example: Tetraploid emmer wheat was domesticated in Turkey (its primary center), but its current center of maximum diversity is in the Mediterranean basin (its secondary center) because it was sympatric with highly diverse wild populations there.
Summary of Key Differences
| Feature | Primary Center | Secondary Center |
|---|---|---|
| Status | Original site of evolution from wild ancestors. | Site of diversity far from the origin. |
| Setting | Uncultivated (mountains, forests). | Cultivated areas. |
| Allele Type | Predominantly dominant alleles. | Predominantly recessive alleles. |
| Traits | Mostly wild characters. | Mostly desirable/domesticated characters. |
| Selection | Natural selection. | Natural and artificial selection. |
17. How do modern genetic studies challenge Vavilov’s theories on origin?
Modern genetic studies, particularly molecular and population genetics, have significantly refined and, in several key areas, directly challenged Nikolai Vavilov’s foundational theories on crop origins. While Vavilov is still considered a “founding pillar” of plant science, the expansion of modern genomic understanding has revealed that the evolutionary history of crops is far more complex than his original models suggested.
The primary challenges to Vavilov’s theories include:
1. The Disconnect Between Diversity and Origin
Vavilov’s central hypothesis was that a crop’s characteristic primary center of diversity is also its center of origin. Modern molecular evidence has demonstrated that this is often not the case.
• The Diversity Paradox: It is now understood that the region with the highest degree of genetic variation may simply be an area where the plant has existed the longest or where human influence and environmental factors have concentrated diversity over time.
• Examples: Crops like Maize and Tomato have centers of maximum genetic diversity that are geographically distinct from their actual centers of origin.
2. The Impact of Subsequent Gene Flow
Molecular studies suggest that gene flow from ancestors after the initial domestication event is the principal cause of the discrepancy between centers of origin and diversity.
• Shifting Centers: If a crop is domesticated from a peripheral population of its wild ancestor, it initially starts with low diversity. However, if that crop later spreads and becomes sympatric (occupying the same area) with highly diverse populations of its ancestor, gene flow can dramatically increase the crop’s diversity in that new region.
• Wheat Examples:
◦ Emmer Wheat: Molecular studies place its origin in the Diyarbakir region of southeastern Turkey, but its center of highest molecular diversity is in the Mediterranean basin.
◦ Common Wheat: Its center of molecular diversity is in Turkey, but its putative origin is in Transcaucasia and northwestern Iran.
3. Rejection of Allele Frequency Patterns
Vavilov proposed that primary centers are characterized by a high frequency of dominant alleles, while recessive alleles increase toward the periphery. Modern genetics has found this view unacceptable and not supported by recent scientific knowledge.
4. Multiple Origins vs. Single Centers
Vavilov generally sought a single point of origin for each crop. Modern research—aided by Carbon-14 dating and molecular markers—indicates that many species had multiple independent centers of origin.
• Common Bean (Phaseolus vulgaris): Modern studies suggest at least two distinct regions of domestication: one in Central America (small-seeded varieties) and one in the Andean region of South America (large-seeded varieties).
5. Geographic Scope and “Non-centers”
Vavilov believed domestication occurred almost exclusively in tropical and subtropical mountainous regions. Modern evidence has challenged this by showing:
• Plains as Origins: Recent findings suggest that plains also served as significant centers of origin for many cultivated plants.
• Non-centers: Scientist Jack Harlan proposed that some crops originated in broad “non-centers”—massive geographical areas where domestication was dispersed over 5,000 to 10,000 kilometers rather than focused in a narrow mountainous “center”.
6. Omitted Regions
Finally, Vavilov’s global surveys were limited by the logistics of his time. He did not adequately cover the vast genetic wealth of Africa or Australia, both of which are now recognized as containing critical genetic diversity for various crop plants.
18. Why do diversity centers often shift away from original sites?
Based on the sources, the shift of a crop’s center of diversity away from its original site (center of origin) is primarily driven by gene flow from ancestral populations and the geographic circumstances of the crop’s initial domestication. This phenomenon, sometimes referred to as a “diversity paradox,” occurs when a crop exhibits higher genetic variation in a region far from where it first appeared.
The key factors causing these shifts include:
1. Subsequent Gene Flow from Ancestors
The primary cause of disagreement between centers of origin and diversity is gene flow from an ancestor occurring after the crop has already originated.
• Introgression: As a domesticated crop spreads geographically, it may come into contact (sympatry) with diverse populations of its wild or weedy progenitors.
• Genetic Enrichment: Continuous gene flow (introgression) from these wild relatives into the crop increases the crop’s genetic variation in that new region, effectively “moving” the center of maximum diversity to where the most diverse ancestral populations reside.
2. Domestication in Peripheral Populations
Diversity centers often shift when a crop is originally domesticated in a peripheral or specialized population of its ancestor.
• Initial Low Diversity: Because peripheral populations often have lower genetic variation than the ancestor’s core population, the newly domesticated crop begins with a narrow genetic base.
• Secondary Diversification: As the crop later migrates into the “center of diversity” of its wild progenitor, it absorbs the existing genetic wealth of that region through hybridization, causing its own diversity center to shift away from its Turkish or Iranian site of origin.
3. Human and Environmental Influence
Diversity may also increase away from the center of origin due to human activities and environmental factors.
• Secondary Centers: Vavilov recognized “secondary centers of origin” where crops developed vast arrays of new variants far from their original homes due to human migration, crossing of different species, and intense natural or artificial selection.
• Adaptation: Over long periods, the accumulation of mutations and recombinations in new environments—aided by human selection for desirable traits—can result in a higher number of cultivars and gene variants than are found at the original site.
Notable Examples from the Sources
• Emmer Wheat: Domesticated in the Diyarbakir region of southeastern Turkey, its center of diversity is now in the Mediterranean. This shift occurred because it was sympatric with the most diverse wild emmer populations in the eastern Mediterranean, picking up diversity through gene flow.
• Common Wheat (Triticum aestivum): While its putative origin is in Transcaucasia and northwestern Iran, its center of molecular diversity is in Turkey. Prolonged sympatry with domesticated and wild tetraploid wheats in Turkey provided a continuous source of new genetic material that shifted its diversity center westward.
• Maize and Tomato: These are cited as classic examples where the known centers of origin and centers of highest genetic diversity currently do not coincide.
19. What are megadiversity centers and which countries have them?
Based on the sources, megadiversity centers (also referred to as megadiversity countries) are defined as a group of 25 selected countries that possess a significant percentage of the Earth’s total species diversity.
Key Characteristics of Megadiversity Centers
• Criteria for Selection: These countries are recognized based on two primary measures of biodiversity: species richness (the total number of species in an area) and endemism (species that are unique to a specific geographic location).
• Geographic Location: Most megadiversity centers are situated in tropical and subtropical regions.
• Global Impact: While they represent a small number of nations, they harbor the majority of the world’s flora and fauna; for example, tropical moist forests alone, which are prevalent in many of these countries, may contain as much as 90% of all known plant species.
Countries and Regions Identified as Megadiversity Centers
While the sources do not provide an exhaustive list of all 25 countries in a single table, they identify many of them within the context of global biodiversity hotspots and centers of origin:
• Asia: India (Hindustan/Himalayan Center), China (East Asian Center), Indonesia, Malaysia, Thailand, and the Philippines.
• North and Central America: Mexico (Central American Center), Costa Rica, Guatemala, and Honduras.
• South America: Brazil, Colombia, Peru, Ecuador, and Bolivia.
• Africa: Ethiopia (Abyssinian Center), Madagascar, and regions of South Africa (such as the Cape Region).
• Oceania: Australia and the Pacific Islands.
Related Concept: Megagene Centers
The sources also mention a related classification called megagene centers, proposed by Zhukovsky. This framework identifies 12 broad geographic regions where the world’s global genetic wealth—including primitive cultivars and their wild and weedy relatives—is distributed. This model expanded upon Nikolai Vavilov’s original eight centers of origin to include:
• Australia.
• The entirety of Africa.
• Siberia.
Preserving these megadiversity centers is considered a high priority because they are the “foundational pillars” of global food security and contain the essential genetic material needed for future crop improvement.
20. Explain the gene pool concept and its breeding levels.
The gene pool concept, also referred to as germplasm or plant genetic resources (PGR), represents the sum total of all alleles influencing the collective hereditary genetic wealth that a crop has acquired over millions of years. Proposed by Harlan and de Wet in 1971, this concept serves as a practical guide for breeders to determine the best strategies for moving genes between related, closely related, and unrelated species.
Instead of focusing on traditional taxonomy, the gene pool concept classifies genetic resources into four levels based on the ease of hybridization and gene transfer.
1. Primary Gene Pool (GP-1)
The primary gene pool is equivalent to the biological species itself.
• Composition: It includes the crop species, cultivated races, landraces, and its wild or weedy spontaneous races.
• Breeding Characteristics: Crossing and gene transfer are easy; hybrids are generally fertile with normal chromosome pairing and gene segregation.
• Usage: Most plant breeders work almost exclusively within this pool to develop commercial cultivars because it avoids the “undesirable linkage blocks” often found in wild relatives.
2. Secondary Gene Pool (GP-2)
This pool includes species that are related to the crop but possess partial barriers to reproduction.
• Composition: It typically includes related species within the same genus (and occasionally outside the genus).
• Breeding Characteristics: Gene transfer is possible but difficult. Hybrids often show some degree of sterility, chromosomes pair poorly during meiosis, and the resulting F1 plants may be weak or difficult to grow to maturity.
• Usage: Recovering desired types in advanced generations is challenging due to the inherent sterility of the hybrids.
3. Tertiary Gene Pool (GP-3)
The tertiary gene pool contains distant relatives where natural crossing is extremely restricted.
• Composition: This group generally includes distant relatives in other genera.
• Breeding Characteristics: Hybridization is either impossible or results in lethal or completely sterile hybrids.
• Breeding Techniques: To obtain successful hybrids, breeders must use drastic techniques such as embryo rescue, chromosome doubling, or bridging species (a third species that facilitates germplasm exchange between the crop and the tertiary relative).
4. Quaternary Gene Pool (GP-4)
Modern biotechnological advances have added a fourth level that bypasses sexual compatibility entirely.
• Breeding Characteristics: This pool is defined by plant transformation techniques (genetic engineering), which allow breeders to move genes across different species, genera, and even different types of organisms.
• Composition: It consists of synthetic strains with DNA or RNA sequences that do not occur in nature within that specific species.
Strategic Importance
The gene pool concept is not static; as new technologies for manipulating genomes are developed, species can move between pools. For example, a species once considered in the tertiary pool may be moved to the secondary pool if new “bridging” or “rescue” techniques make gene transfer more feasible. Breeders utilize these pools to find essential traits—such as pest and disease resistance or stress tolerance—that may no longer be present in modern, elite cultivars.
21. How do breeders use different gene pools for crop improvement?
Breeders use the gene pool concept as a practical guide to determine the most effective strategies for moving desired traits between related, closely related, and unrelated species. Based on the ease of hybridization and gene transfer, breeders categorize genetic resources into four distinct levels to facilitate crop improvement.
1. Primary Gene Pool (GP-1): The Preferred Source
The primary gene pool is equivalent to the biological species and includes cultivated varieties, landraces, and their immediate wild or weedy ancestors.
• Usage: Breeders work exclusively within this pool for the vast majority of commercial cultivar development.
• Mechanism: Crossing and gene transfer are easy; hybrids are generally fertile with normal chromosome pairing and gene segregation.
• Advantage: It is the primary source of variation because it avoids “undesirable linkage blocks” and negative interactions often found when using wild relatives.
2. Secondary Gene Pool (GP-2): For Hard-to-Reach Traits
This pool includes related species, typically within the same genus, where gene transfer is possible but difficult.
• Usage: Breeders turn to this pool when a desired trait—such as specific stress tolerance—is missing in GP-1.
• Challenges: Hybrids (F1 plants) are often weak, exhibit poor chromosome pairing during meiosis, and tend to be sterile.
• Breeding Effort: Because of this partial sterility, recovering desired types in advanced generations is challenging and requires significant effort to eliminate “linkage drag” from the wild parent.
3. Tertiary Gene Pool (GP-3): Drastic Breeding Techniques
The tertiary gene pool consists of distant relatives, often in other genera, where natural hybridization is extremely restricted.
• Usage: It is used as a last resort to access extreme resistance traits.
• Advanced Techniques: To obtain successful hybrids, breeders must use “drastic techniques” such as embryo rescue, chromosome doubling to restore fertility, or bridging species.
• Example: Breeders successfully used a translocation to transfer a disease resistance cluster from rye (a tertiary relative) to bread wheat, which was subsequently planted on millions of hectares.
4. Quaternary Gene Pool (GP-4): Bypassing Sexual Barriers
Modern biotechnology has introduced a fourth level that bypasses traditional sexual cross-compatibility.
• Mechanism: Breeders use plant transformation techniques (genetic engineering) to move specific DNA or RNA sequences across different species, genera, or even different organisms entirely.
• Usage: This allows for the creation of synthetic strains with traits that do not occur naturally within that species.
Integrated Breeding Strategies
• Prebreeding (Germplasm Enhancement): This is the systematic process of integrating genes from non-elite sources (GP-2 and GP-3) into adapted genotypes. Breeders often use the cultivated type as the female parent and the wild relative as the male, followed by repeated backcrossing to reduce undesirable wild traits while retaining the target gene.
• Marker-Assisted Selection (MAS): Modern breeders use molecular markers to track the inheritance of these “wild” genes more effectively during the prebreeding process.
• Dynamic Nature: Gene pools are not static; as new technologies like gene editing or embryo rescue improve, species once considered “tertiary” may move into the “secondary” gene pool.
Breeders continuously evaluate these pools to find “raw ingredients”—such as resistance to emerging biotypes of pests or new environmental challenges—to ensure long-term agricultural sustainability.
References
Dvorak, J., Luo, M.-C., & Akhunov, E. D. (2010). N.I. Vavilov’s Theory of Centres of Diversity in the Light of Current Understanding of Wheat Diversity, Domestication and Evolution. http://staden.sourceforge.net/
Engels, J. M. M., Ebert, A. W., Thormann, I., & De Vicente, M. C. (2006). Centres of crop diversity and/or origin, genetically modified crops and implications for plant genetic resources conservation. In Genetic Resources and Crop Evolution (Vol. 53, Number 8, pp. 1675–1688). https://doi.org/10.1007/s10722-005-1215-y
Hummer, K. E., & Hancock, J. F. (2015). Vavilovian Centers of Plant Diversity: Implications and Impacts. HortScience, 50(6). https://journals.ashs.org
Kumar, C., Kotra, V., Kumar, N., Singh, K., & Singh, A. K. (2023). Biodiversity and bioresources: impact of biodiversity loss on agricultural sustainability. In Biodiversity and Bioeconomy: Status Quo, Challenges, and Opportunities (pp. 165–198). Elsevier. https://doi.org/10.1016/B978-0-323-95482-2.00008-0
Singh, D. P., Singh, A. K., & Singh, A. (2021). Plant genetic resources. In Plant Breeding and Cultivar Development (pp. 129–157). Elsevier. https://doi.org/10.1016/b978-0-12-817563-7.00009-x
Related posts:
 ढलौटे फोहोरको बायोचार र Trichoderma afroharzianum सँग मिलेर टमाटरमा हुने जराको रोग नियन्त्रण
ढलौटे फोहोरको बायोचार र Trichoderma afroharzianum सँग मिलेर टमाटरमा हुने जराको रोग नियन्त्रण
 The Evolutionary Genomics of Crop Plants: From Ancestral Polyploidy to Modern Pangenomes
The Evolutionary Genomics of Crop Plants: From Ancestral Polyploidy to Modern Pangenomes
 Global Strategies for Crop Germplasm Conservation: Integrating Farmer-Based In Situ Methods with Biotechnological Innovation
Global Strategies for Crop Germplasm Conservation: Integrating Farmer-Based In Situ Methods with Biotechnological Innovation
 How Wild Soybeans Use Microbes to Thrive in Salty Soils: Lessons from the Yellow River Delta
How Wild Soybeans Use Microbes to Thrive in Salty Soils: Lessons from the Yellow River Delta
 Biochar-Enhanced Soil Treatment Cuts Antibiotic Resistance Genes by Over 59%
Biochar-Enhanced Soil Treatment Cuts Antibiotic Resistance Genes by Over 59%

