1. Introduction: The Strategic Importance of Mating Systems
In reproductive ecology, the mating system is defined as the distribution of “who mates with whom and how often” within a sexually reproducing population. This distribution is a primary determinant of genetic diversity and a central modulator of sexual selection. By dictates of genetic transmission, a species’ mating strategy influences the distribution of alleles and the efficacy of natural selection, shaping everything from the morphological intricacies of floral architecture to the behavioral complexities of animal courtship.

These notes provide a synthesis of current research on the evolution of mating strategies, bridging the historical divide between botanical and zoological frameworks. We will evaluate the evolutionary “logic” of inbreeding, the biological taxes imposed by inbreeding depression, and the mechanisms—ranging from temporal shifts to morphological adaptations—that organisms employ to navigate these trade-offs. Finally, we address the strategic necessity of understanding anthropogenic shifts in pollination biology, as human activity increasingly dictates the evolutionary trajectory of global flora.

2. Conceptual Framework: Defining Mating Systems Across Taxa
A coherent definition of mating systems is essential to integrate research across the plant-animal divide. Historically, plant biology has focused on the “selfing rate” (s)—the proportion of progeny produced via self-fertilization—while animal research has centered on pair bonds and mating structures (e.g., polygyny, monogamy) in gonochoric populations.
This gap is bridged by shifting the focus to “patterns of fertilization.” By analyzing fertilization events rather than physical copulation, we can apply a unified evolutionary framework to disparate taxa. This allows us to treat a wind-pollinated tree and a broadcast-spawning marine invertebrate as having analogous mating dynamics, where gamete release into the environment replaces direct sexual encounters.
| Dimension | Hermaphroditic Plants | Gonochoric Animals |
|---|---|---|
| Primary Focus of Research | Selfing (s) vs. Outcrossing (t) rates | Pair bonds and mating structures |
| Typical Mating Risk | High inbreeding risk due to proximity | Lower risk due to pre-mating dispersal |
| Fundamental Challenge | Decision to self-fertilize vs. outcross | Competition for mate and resource access |
Note: These dimensions represent generalizations used to highlight different research histories; however, “patterns of fertilization” provides the specific conceptual tool required for a cohesive cross-taxa analysis.

3. The Advantages of Inbreeding: Transmission and Assurance
Evolutionary theory reveals that inbreeding follows a specific biological logic, moving beyond the perception of it being inherently deleterious.
The 1.5-fold “Transmission Advantage” (Fisher’s Insight)
Fisher (1941) identified that an allele promoting selfing in a hermaphroditic population possesses a 50% (1.5-fold) transmission advantage over outcrossing alleles at autosomal loci:
• Dual Contribution: A selfing individual contributes both male and female gametes to its own offspring, ensuring two copies of its genome are transmitted.
• Siring Opportunities: While selfing its own ovules, the individual can often still contribute pollen to the outcrossing pool, effectively siring additional offspring.
• Allele Efficiency: This dual path allows selfing alleles to propagate more rapidly than those restricted to outcrossing.

Reproductive Assurance (RA) and Baker’s Law
Self-fertilization provides a demographic safety net known as Reproductive Assurance. In scenarios of mate or pollinator scarcity—common during the colonization of islands or range margins—the capacity to self-fertilize prevents extinction from reproductive failure. This is the foundation of Baker’s Law, which is described as:
“…the demographic sieve on preexisting variation in the mating system that is present during long-distance dispersal, leading to colonization of new patches.”
Lineages capable of self-fertilization are thus prioritized by this “sieve,” making them superior colonizers frequently found at high latitudes or on oceanic islands.
4. The Costs of Inbreeding: Depression and Discounting
Genetic and demographic costs balance the advantages of inbreeding. The realized mating system represents a negotiation between the benefits of transmission and the biological “taxes” of homozygosity.

Inbreeding Depression (ID) and the 0.5 Threshold
Inbreeding depression is the reduction in fitness of inbred offspring due to the expression of deleterious recessive mutations. Following the Lande & Schemske (1985) model, outcrossing is favored only when ID exceeds 0.5. If the fitness of selfed offspring is less than half that of outcrossed offspring, selection maintains outcrossing; otherwise, complete selfing is the predicted stable state.
Pollen Discounting and Effective Population Size (Ne)
Pollen discounting occurs when self-fertilization reduces the amount of pollen available for outcrossing, thereby diminishing the transmission advantage. Furthermore, inbreeding rescales the effective population size (Ne). Using the relationship Ne=N/(1+F), where F is the inbreeding coefficient (FIS), we see that increased inbreeding directly reduces Ne, accelerating genetic drift.
Purging: Drift vs. Active Inbreeding
The rate of inbreeding determines the efficacy of “purging” deleterious mutations:
• “Slow” Inbreeding: Characteristic of biparental inbreeding in animals. High Ne is maintained, allowing selection to efficiently remove mutations of any dominance level.
• “Fast” Inbreeding: Characteristic of selfing in plants. While it exposes mutations quickly, the massive reduction in Ne increases drift. Consequently, drift in small populations tends to purge only highly recessive mutations, while active inbreeding can potentially remove alleles regardless of dominance, provided the population avoids “mutational meltdown.”
5. Mechanisms of Mating Strategy and Inbreeding Avoidance
Organisms utilize anatomical, temporal, and physiological mechanisms to determine their realized mating system.
• Temporal/Spatial Strategies:
◦ Prior Selfing: Occurs before floral opening.
◦ Competing Selfing: Simultaneous deposition of self and outcross pollen.
◦ Delayed Selfing: Selfing occurs only during flower senescence if outcrossing failed, providing RA with minimal cost.
• Morphological Strategies and Bet-Hedging:
◦ Geitonogamy: Transfer of pollen between flowers on the same plant. Larger floral displays increase this risk and its associated pollen discounting.

◦ Cleistogamy: Weissman et al. (2024) identify closed flowers as a bet-hedging strategy in unstable biomes, ensuring reproduction when environmental conditions are unpredictable.

• Avoidance Systems: Self-Incompatibility (SI) and Dioecy (separate sexes) are primary barriers to inbreeding.

The “Dead-End Hypothesis”
The transition from outcrossing to selfing is generally unidirectional. Evidence from taxa such as Linanthus (section Leptosiphon), Caenorhabditis nematodes, and social spiders (genus Stegodyphus) supports the hypothesis that while selfing provides short-term benefits, the resulting loss of genetic diversity and reduced efficacy of selection leads to higher extinction rates for selfing lineages.
1. Reduced Adaptive Potential: The loss of genetic diversity and reduced Ne limit responses to environmental shifts or novel pathogens.
2. Unidirectional Evolution: Transitions to selfing are often permanent. Once mechanisms like self-incompatibility are lost, reversion to outcrossing is nearly impossible.
3. Accumulation of Mutation Load: The combination of increased drift and reduced selection leads to a “mutational meltdown.”
Phylogenetic signals support this: selfing lineages frequently appear at the “tips” of phylogenetic trees in both angiosperms and the nematode genus Caenorhabditis (Goldberg et al., 2010; Cutter et al., 2019), indicating they do not persist long enough to diversify.

6. Anthropogenic Impact: Mating Systems in a Changing World
Human activity is fundamentally altering pollination landscapes, necessitating a shift toward self-fertilization for many species.
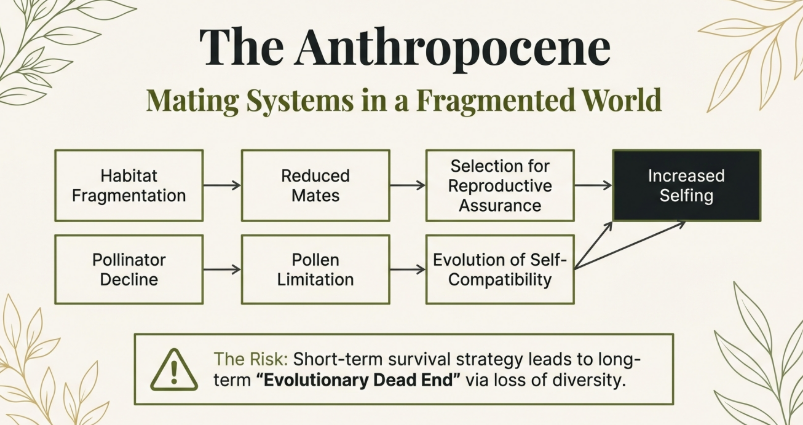
Habitat Fragmentation and Outcross Pollen Limitation (PLx)
A meta-analysis demonstrates a significant reduction in the outcrossing rate (t) in disturbed habitats. Fragmentation reduces Mate Availability and Pollinator Availability, intensifying PLx. While fragmentation reduces both, the selective pressure for RA specifically targets the lack of outcross pollen, favoring autofertility.
Glossary of Pollination Parameters
• Autofertility (AF): The capacity to produce seed via autonomous self-pollination in the absence of pollinators.
• Pollen Limitation (PLx): The degree to which seed production is restricted specifically by a lack of outcross pollen.
• Emasculation (FEN): A parameter representing the seed production of emasculated flowers; used to isolate the outcrossing rate in natural environments.
7. Modern Environmental Risks and Conservation Implications
Anthropogenic disturbance serves as an unplanned experiment in mating-system evolution, often forcing outcrossing species toward selfing.
Human-Induced Shifts and Scale
A meta-analysis of 22 studies involving 27 plant species demonstrates that habitat modification significantly reduces the proportion of seeds outcrossed (t). This shift is particularly severe in self-compatible species where pollinators or mates are scarce.
Life-History Nuance: Annuals vs. Perennials
While annuals may adopt selfing for immediate survival, long-lived perennials (e.g., trees) exhibit different risks. Though they can withstand poor breeding seasons better, their large floral displays make them more prone to geitonogamy (selfing via pollinators visiting multiple flowers on the same plant).
The Conflict of Limitation
Conservation strategies must distinguish between:
• Pollinator Limitation: Lack of vectors preventing pollen transfer.
• Mate Limitation: Lack of conspecifics, meaning even active pollinators transfer only self or related pollen.
8. Future Research Directions and Conclusion
While self-fertilization offers immediate reproductive assurance, it frequently leads to evolutionary failure. To mitigate extinction risks in vulnerable populations, we must address the following research gaps:
• Dispersal-Inbreeding Feedbacks: Investigating how the loss of pre-mating dispersal (philopatry) accelerates transitions to sociality and obligate inbreeding in arthropods and other animals.
• Adaptive vs. Tolerated Selfing: Testing if selfing in natural populations is a truly adaptive strategy or a non-adaptive byproduct of small population size and drift.
• Theoretical Syntheses: Developing explicit models that compare the long-term genomic trajectories of selfing versus biparental (sib-mating) systems to refine extinction risk predictions.
Bridging the plant-animal divide in mating-system research is essential for developing robust conservation strategies that preserve not just current numbers, but the evolutionary potential of species.
A cohesive approach to mating-system evolution is no longer optional; it is a requirement for understanding the persistence of sexually reproducing organisms in the Anthropocene.
Image Summary





Questions/Answers
1. How do plants co-evolve with pollinators to ensure successful mating?
Plants co-evolve with pollinators through a process called pollinator-mediated selection, where the two groups exert reciprocal selective pressure on each other to maximize reproductive success and fitness. This co-evolution results in specialized floral traits and behaviors designed to ensure the efficient transfer of pollen while managing the trade-offs between self-fertilization and outcrossing.
Plants ensure successful mating through the following co-evolved strategies:
1. Pollination Syndromes
Plants accumulate suites of related traits, known as pollination syndromes, that match the sensory and physical requirements of specific pollinators.
• Visual and Olfactory Attractants: Plants use specific colors and scents to signal rewards. For example, bee-pollinated flowers are often yellow or blue with ultraviolet nectar guides, while moth-pollinated flowers are typically white, night-opening, and emit sweet scents in the evening.
• Nutritional Rewards: Plants produce nectar (sugar-dominated for bees, copious and dilute for birds) and edible pollen to encourage repeat visits.
• Mechanical Matching: Striking examples of co-evolution include the development of long floral corollas that match the long beaks or proboscii of specialized pollinators like hummingbirds or moths.
2. Structural Adaptations (Floral Design)
Plants utilize spatial and temporal separation of sex organs to reduce sexual interference—where a plant’s own male and female functions conflict—and to promote outcrossing.
• Herkogamy: This is the spatial separation of pollen presentation (anthers) and receipt (stigma).
◦ Reciprocal Herkogamy: In polymorphisms like heterostyly (different style lengths) and enantiostyly (mirror-image flower shapes), plants ensure that pollen is placed on a specific part of a pollinator’s body that corresponds exactly to the position of the stigma in a different floral morph, promoting cross-pollen transfer.
• Dichogamy: This involves the temporal separation of sex functions. In protandry, pollen is released before the stigma is receptive; in protogyny, the reverse occurs. This prevents the plant from self-pollinating and forces the pollinator to move between flowers in different phases.
3. Highly Specialized “Trickery”
Some plants, particularly orchids, have evolved extreme specializations that ensure mating without offering nutritional rewards.
• Mimicry: Certain orchids mimic the shape and scent (pheromones) of female insects to attract males into attempting to “mate” with the flower, thereby attaching pollen clusters called pollinia to the insect.
• Traps: Some fly-pollinated or beetle-pollinated flowers use traps to hold pollinators for extended periods, ensuring they come into contact with pollen before they leave.
4. Strategic Floral Displays
The arrangement and number of open flowers on a plant—the floral display—are co-evolved to attract pollinators while minimizing the risks of geitonogamy (selfing between flowers on the same plant).
• Geitonogamy and Pollen Discounting: Large displays attract more pollinators but increase the chance they will visit multiple flowers on the same plant, leading to self-pollination. This causes pollen discounting, where pollen that could have been exported to other plants is “wasted” on selfing, reducing paternal fitness.
• Synchrony vs. Asynchrony: To avoid “stigma gunking” or receiving pollen from the wrong species, plants may flower at different times (niche partitioning) or synchronously to overwhelm pollinators with a visual signal.
5. Reproductive Assurance
Co-evolution also accounts for scenarios where pollinators are unreliable.
• Delayed Selfing: Some plants have evolved mechanisms to self-pollinate only after opportunities for outcrossing have passed, ensuring they produce seeds even if no pollinators visit.
• Self-Incompatibility (SI): To prevent the negative effects of inbreeding depression, many plants have evolved physiological systems to recognize and reject their own pollen, forcing successful mating to occur only with unrelated individuals.
In summary, plants do not simply “attract” pollinators; they use a complex suite of co-evolved morphological and phenological strategies to manipulate pollinator behavior, ensuring that pollen is not only moved but moved to the right destination to produce high-quality, outcrossed offspring.
2. What are the trade-offs between self-fertilization and outcrossing strategies?
The choice between self-fertilization (selfing) and outcrossing represents a fundamental evolutionary conflict in plants, involving complex genetic, demographic, and resource-based trade-offs.
1. Genetic Trade-offs
• The Transmission Advantage: The most significant benefit of selfing is the automatic gene transmission advantage (Fisher’s 1.5-fold advantage). A selfing individual transmits three copies of its genes to the next generation (two through its own seeds and one through its pollen fertilizing others) for every two copies transmitted by an outcrosser.
• Inbreeding Depression: The primary cost of selfing is inbreeding depression, which refers to the reduced survival and fertility of inbred offspring due to the expression of deleterious recessive mutations. If the fitness cost of inbreeding depression exceeds 50%, outcrossing is typically favored; otherwise, selfing alleles tend to spread.
• Purging vs. Genetic Dead-End: In the long term, selfing can purge deleterious alleles by exposing them in a homozygous state. However, selfing also reduces effective population size (Ne) and recombination rates, making a population more susceptible to genetic drift and less able to adapt to environmental changes. Consequently, selfing is often viewed as an evolutionary dead-end that increases extinction risk.
2. Demographic and Ecological Trade-offs
• Reproductive Assurance: Selfing provides reproductive assurance, allowing plants to set seed when pollinators or potential mates are scarce. This is a critical advantage for weeds, colonizing species, and plants at the margins of their geographic range (Baker’s Law).
• Pollination Failure: Outcrossing relies on external vectors (wind, water, or animals). If these services are unreliable, outcrossers may suffer from pollen limitation, resulting in unfertilized ovules and reduced fitness.
• The Cost of Attraction: Outcrossing strategies require significant resource investment in floral design and display—such as nectar, scent, and showy petals—to attract pollinators. Over time, selfing lineages often evolve a “selfing syndrome,” characterized by smaller, less costly flowers.
3. Resource and Mating Efficiency Trade-offs
• Pollen Discounting: This is a cost of selfing where the pollen used for self-fertilization is no longer available to be exported to other plants, thereby reducing the individual’s paternal fitness (male outcross success).
• Seed Discounting: Self-fertilization may use up ovules or resources that could have otherwise produced higher-quality outcrossed seeds.
• Geitonogamy (Maladaptive Selfing): In plants with large floral displays meant to attract pollinators, geitonogamy (pollen transfer between flowers on the same plant) often occurs. This is frequently maladaptive because it incurs the costs of inbreeding depression and pollen discounting without providing the reproductive assurance benefits of autonomous selfing.
4. Summary of Trade-offs
| Feature | Self-Fertilization (Selfing) | Outcrossing |
|---|---|---|
| Gene Transmission | High (1.5x advantage) | Standard |
| Offspring Fitness | Low (Inbreeding depression) | High (Heterosis) |
| Mate Availability | Not required (Reproductive assurance) | Obligate need for mates |
| Resource Cost | Low (Small flowers, no nectar) | High (Attraction strategies) |
| Adaptive Potential | Low (Reduced Ne and recombination) | High (Increased genetic diversity) |
3. Explain how human environmental disturbance impacts plant mating and diversity.
Human environmental disturbance, including habitat fragmentation, selective logging, and climate change, significantly alters plant mating patterns and diminishes genetic diversity. These disturbances primarily impact plant reproduction by reducing outcrossing rates—the proportion of seeds produced through cross-fertilization with unrelated individuals—while increasing self-fertilization.
Impacts on Plant Mating Systems
Human activities disrupt the essential interactions between plants and their pollinators through several mechanisms:
• Reduction in Outcrossing: A meta-analysis of 22 studies shows a significant reduction in outcrossing (t) in disturbed habitats compared to undisturbed ones. For example, the tropical tree Shorea curtisii exhibited near-complete outcrossing in undisturbed forests, but this dropped to 50% in selectively logged areas.
• Reduced Pollinator and Mate Availability: Disturbance often leads to lower pollinator abundance and creates smaller, sparser plant populations. When conspecific plant density is low, pollinators may carry less conspecific pollen or deposit higher amounts of “contaminating” heterospecific pollen from other species, interfering with fertilization.
• Evolution of Reproductive Assurance: Chronic pollen limitation caused by disturbance selects for selfing as a mechanism of reproductive assurance (RA). Plants may evolve floral traits that facilitate autonomous self-pollination (autofertility) to ensure seed production when pollinators or mates are scarce.
• Shift to Selfing Syndromes: In severely disturbed habitats with a chronic lack of pollinators and mates, populations may eventually evolve a “selfing syndrome,” characterized by reduced investment in attractive structures (like large petals), less nectar production, and shorter floral lifespans.
Impacts on Genetic Diversity
The shift from outcrossing to inbreeding has profound consequences for a population’s genetic health and evolutionary trajectory:
• Increased Inbreeding and Genetic Load: Most plant species harbor deleterious recessive mutations; increased selfing expresses this “genetic load,” leading to inbreeding depression, which reduces progeny vigor, survival, and fertility.
• Reduced Effective Population Size (Ne): Inbreeding effectively halves the effective population size compared to outcrossing populations of the same census size. This reduction accelerates genetic drift, causing a more rapid loss of neutral genetic diversity.
• Reduced Adaptive Potential: Because selfing reduces recombination rates and genetic variation, disturbed populations may have a limited ability to adapt to further environmental changes.
• Population Differentiation and Extinction Risk: Habitat fragmentation increases population differentiation (FST) and restricts gene flow. Over time, the accumulation of mildly deleterious mutations and the loss of diversity may lead to population deterioration or extinction, a concept often called the “evolutionary dead-end” hypothesis.
In summary, human environmental disturbance forces many plant species toward increased self-fertilization to ensure immediate survival (reproductive assurance), but this shift comes at the cost of reduced genetic diversity and long-term fitness, potentially leading to local extirpations.
4. How do plants resolve the conflict between display size and self-pollination?
Plants resolve the conflict between the need for large floral displays to attract pollinators and the resulting risk of geitonogamy (self-pollination between flowers on the same plant) through various structural, temporal, and phenological strategies. While large displays increase visual and olfactory signals, they also encourage pollinators to remain on a single plant longer, which increases self-fertilization and leads to mating costs like inbreeding depression and pollen discounting.
Plants utilize the following mechanisms to decouple the benefits of attraction from the costs of selfing:
1. Temporal and Spatial Separation (Dichogamy and Herkogamy)
Plants use precise floral designs to minimize sexual interference and geitonogamy:
• Synchronized Protandry: In vertical inflorescences (like racemes), flowers often mature sequentially from bottom to top. Because pollinators like bees typically forage from the bottom of an inflorescence upward, plants ensure they receive outcross pollen on lower female-phase flowers first, then pick up fresh pollen from upper male-phase flowers before flying to a different plant.
• Reciprocal Herkogamy: Systems like heterostyly (different style lengths) and enantiostyly (mirror-image flowers) ensure that pollen is deposited and received on specific parts of a pollinator’s body. This spatial segregation reduces the chance of self-pollen being deposited on a plant’s own stigmas even when multiple flowers are visited.
2. Phenological Adjustments (Individual Flowering Strategies)
Plants may adjust their flowering phenology—the timing and rate at which flowers open—to balance attraction and mating quality:
• Extended Flowering vs. Mass Blooming: To avoid the “geitonogamy constraint” associated with mass blooming, many plants evolve longer flowering periods with fewer flowers open simultaneously each day. This strategy favors outcrossing by reducing the number of consecutive visits a pollinator makes to flowers on the same individual.
• Niche Partitioning: Some species may flower asynchronously with competitors to ensure more reliable visits from pollinators, reducing the “waste” of pollen during simultaneous multi-species blooms.
3. Sexual System Transitions
In cases where large size or extensive clonal growth makes geitonogamy inevitable and highly damaging, plants may evolve entirely different sexual systems:
• Monoecy: By producing separate male and female flowers on the same plant, plants can spatially or temporally segregate sex functions more effectively than in hermaphroditic flowers.
• Dioecy (Separate Sexes): Dioecy is considered the ultimate evolutionary solution to the geitonogamy constraint. By being exclusively male or female, individuals can produce massive floral displays to maximize pollinator attraction without any risk of self-fertilization.
4. Clonal Architecture
For clonal plants, where multiple “apparent individuals” (ramets) are genetically identical, geitonogamy is a major risk. Some plants resolve this through:
• Fragmentary Growth: Highly clonal aquatics may utilize weak stolon connections that break, allowing daughter rosettes to drift and mix spatially with different genotypes.
• Guerilla vs. Phalanx Strategies: Plants may evolve “guerilla” growth forms, where clones are intermingled with other genotypes, rather than “phalanx” forms where identical clones are clumped together, thereby increasing the likelihood of outcrossing during pollinator movement.
In summary, plants manage the “showiness” required for reproductive success by employing floral designs that manipulate pollinator movement and phenologies that limit daily flower availability, ensuring that pollen is exported to other individuals rather than being lost to self-pollination.
5. What are the genetic costs of geitonogamy and pollen discounting?
In plant mating systems, geitonogamy and pollen discounting represent significant genetic costs that can undermine the evolutionary advantages of self-fertilization. Together with inbreeding depression, these factors are the primary forces selecting against selfing and promoting the maintenance of outcrossing.
1. The Genetic Costs of Geitonogamy
Geitonogamy occurs when pollinators move between different flowers on the same individual plant. While physically involving multiple flowers, it is genetically equivalent to selfing. Its primary genetic costs include:
• Inbreeding Depression (Maternal Cost): Because geitonogamy results in selfed offspring, it exposes them to inbreeding depression—the reduction in viability and fertility caused by the expression of deleterious recessive mutations in a homozygous state. This significantly reduces the individual’s fitness as a maternal parent.
• Maladaptive Nature: Unlike autonomous self-pollination within a single flower, which can provide “reproductive assurance” when pollinators are scarce, geitonogamy requires a pollinator to occur. Consequently, it is often viewed as a maladaptive consequence of having large floral displays intended to attract pollinators; the plant pays the “cost” of inbreeding without receiving the benefit of guaranteed seed set in the absence of mates.
• Induction of Pollen Discounting: Geitonogamy is a major driver of pollen discounting because each visit to another flower on the same plant “wastes” pollen that could have been exported elsewhere.
2. The Genetic Costs of Pollen Discounting
Pollen discounting is the reduction in an individual’s outcrossed siring success (paternal fitness) caused by its own self-pollination. Its genetic consequences include:
• Reduced Paternal Fitness: Pollen used for self-fertilization is no longer available for export to other plants in the population. This reduces the individual’s contribution to the next generation as a paternal (male) parent.
• Negating the Transmission Advantage: In theory, selfing alleles have a 1.5-fold “automatic transmission advantage” (Fisher’s advantage), where a selfing individual transmits three copies of its genes for every two transmitted by an outcrosser. High levels of pollen discounting can abolish this advantage entirely; if selfing prevents an individual from siring any outcrossed seeds on other plants, the evolutionary incentive for selfing disappears.
• Evolutionary Stability of Outcrossing: Pollen discounting effectively increases the “cost” of inbreeding. Even if inbreeding depression is relatively low, high pollen discounting can prevent selfing mutants from invading and taking over an outcrossing population.
Summary of Genetic Trade-offs
| Cost Type | Mechanism | Fitness Impact |
|---|---|---|
| Maternal | Inbreeding depression from selfed seeds | Reduced quality of offspring (lower survival/fertility). |
| Paternal | Pollen discounting | Reduced quantity of outcrossed siring opportunities (male success). |
| Evolutionary | Interaction of the above | Negates the 50% gene transmission advantage typically associated with selfing. |
To mitigate these costs, many plants have evolved structural or temporal strategies—such as dichogamy (separating male and female functions in time) or herkogamy (separating them in space)—that reduce geitonogamy and pollen discounting while still allowing for large, attractive floral displays.
6. How does self-incompatibility function as a mate recognition system?
Self-incompatibility (SI) functions as a physiological and chemical mate recognition system that allows plants to distinguish between self and non-self pollen, effectively acting as a filter to prevent inbreeding. Despite its name, it is not strictly a “self-recognition” system but is instead based on molecular interactions between receptors and ligands encoded by specific genes.
The system functions through the following mechanisms:
1. Genetic Basis: The S-Locus
The recognition process is primarily governed by a specific genetic region known as the S-locus (self-incompatibility locus). When a pollen grain reaches the stigmatic surface of a plant, the system checks for a matching allele at this locus. If the allele in the pollen matches an allele in the maternal plant’s pistil, the pollen is recognized as “self” or a matching genotype and is subsequently rejected.
2. Molecular Mechanisms of Recognition
The system relies on proteins with receptors and ligands that recognize one another. There are two primary genetic modes of recognition:
• Gametophytic SI (GSI): In this system, the incompatibility phenotype of the pollen is determined by its own haploid genome. Rejection often involves S-RNases in the style and F-box proteins in the pollen grains. Examples include the Solanaceae and Rosaceae families.
• Sporophytic SI (SSI): Here, the incompatibility is determined by the diploid genotype of the parent plant that produced the pollen. This system is characteristic of the Brassicaceae family.
3. Stages of Rejection
Once a matching allele is recognized, the plant triggers physiological responses to stop the mating process at various stages:
• Germination and Growth: The plant may inhibit pollen germination or pollen-tube growth as the tube attempts to travel down the style.
• Ovarian or Late-Acting SI: In some species, such as Narcissus, recognition and rejection occur even later, within the ovary. This can involve “long-distance” signaling between pollen tubes and the ovary, resulting in embryo sac degeneration following self-pollination.
• Fertilization and Development: If the previous stages are bypassed, the system can still inhibit ovule fertilization or embryo development, ensuring no seeds are produced from a “self” match.
4. Evolutionary Purpose
As a mate recognition system, SI is one of the most significant means of preventing inbreeding depression—the reduced fitness of offspring resulting from the expression of deleterious recessive mutations. By forcing obligate outcrossing, SI promotes the generation of new genotypes and is considered a major factor in the evolutionary success and radiation of angiosperms. While SI is an effective barrier, it can sometimes be “leaky,” and some species exhibit “pseudo-self-fertility” where the system breaks down under specific environmental conditions or successional stages.
7. How do plants purge deleterious mutations through self-fertilization?
Plants purge deleterious mutations through self-fertilization by increasing the frequency of homozygous genotypes, which exposes recessive harmful alleles to the forces of natural selection. This process, often referred to as purging the genetic load, is a critical mechanism in the evolution and stabilization of plant mating systems.
The purging process occurs through several key stages:
1. Increase in Homozygosity
Self-fertilization rapidly increases the frequency of homozygotes within a population compared to random outcrossing. While outcrossing tends to mask deleterious recessive mutations in a heterozygous state, selfing brings these alleles together in pairs.
2. Expression of Recessive Alleles
Many harmful mutations are only expressed when an individual carries two copies of the mutant allele (homozygous recessive). By increasing homozygosity, selfing forces these deleterious recessive phenotypes to be expressed, which would otherwise remain hidden and “protected” from selection in an outcrossing population.
3. Reduced Fitness and Inbreeding Depression
The expression of these deleterious mutations typically leads to inbreeding depression, which manifests as reduced survival probabilities and lower fertility for the affected individuals. Inbreeding depression serves as the “filter” that allows selection to identify unfit individuals.
4. Selective Removal (Purging)
Because individuals with these homozygous deleterious mutations have lower fitness, they are less likely to survive to maturity or produce viable offspring. Natural selection thus eliminates these alleles from the breeding pool. Over many generations, this process lowers the overall frequency of harmful mutations in the population, effectively “purging” the genetic load and reducing the severity of future inbreeding depression.
5. Efficiency and Rate of Purging
The sources highlight that the efficiency of purging depends on the rate of inbreeding:
• “Fast” Inbreeding: Extreme self-fertilization exposes mutations very quickly but also significantly reduces the effective population size (Ne). This reduction in Ne can increase the impact of genetic drift, which may accidentally “fix” mildly deleterious mutations rather than eliminating them.
• “Slow” Inbreeding: Lower rates of inbreeding (such as mating with relatives) may allow for more efficient purging because the effective population size remains larger, allowing natural selection to be more precise in removing disadvantageous alleles while increasing the frequency of advantageous ones.
Long-Term Evolutionary Impact
If a population persists long enough to successfully purge its deleterious mutations, it may evolve low inbreeding depression, which makes a return to an outcrossing strategy highly unlikely. This is because the primary genetic cost of selfing—the fitness reduction of inbred offspring—has been removed. However, the sources also note the “dead-end” hypothesis, which suggests that while purging removes harmful mutations, the overall loss of genetic diversity and reduced recombination in selfing lineages may still lead to higher long-term extinction rates.
8. What is reproductive assurance in plants?
Reproductive assurance (RA) in plants is the increase in seed production afforded by self-pollination when a scarcity of pollinators or potential mates limits outcross pollination. It is considered a primary evolutionary advantage of self-fertilization, ensuring that a plant maintains some level of fitness even when outcrossing services are unreliable, unpredictable, or absent.
According to the sources, reproductive assurance involves several mechanisms, ecological contexts, and evolutionary trade-offs:
1. Mechanisms of Reproductive Assurance
For reproductive assurance to be effective, a plant must be self-compatible and possess mechanisms to facilitate the transfer of its own pollen to its stigmas.
• Autonomous Autogamy: This is spontaneous self-pollination that occurs without the intervention of pollinators. This is the most common form of RA because it directly compensates for a lack of animal visitation.
• Delayed Selfing: Often described as the “best of both worlds,” this occurs after all opportunities for outcrossing have passed, such as when flowers are senescing (wilting). It allows a plant to prioritize high-quality outcrossed seeds but “fails safe” to selfed seeds if outcrossing does not occur.
• Prior Selfing: This occurs before the flower even opens (e.g., cleistogamy or selfing in an unopened bud). This ensures reproduction but reduces outcrossing opportunities and can save resources by eliminating the need for showy floral displays.
• Autofertility (AF): This refers to the capacity of a plant to produce seeds via autonomous selfing when pollinators are excluded (often tested by caging plants). While autofertility indicates the potential for RA, actual reproductive assurance is measured as the actual increase in seed set under natural conditions when outcrossing is limited.
2. Ecological Significance and “Baker’s Law”
Reproductive assurance is a critical strategy for survival in specific ecological scenarios:
• Colonization and Invasions: RA is at the heart of Baker’s Law, which proposes that self-compatible individuals are more likely to successfully establish populations after long-distance dispersal because they do not require a mate or specific pollinators to produce offspring.
• Range Margins and Disturbance: RA is frequently observed at the margins of a species’ geographic range, in small or isolated populations, and in habitats experiencing high human disturbance. In these areas, mates are often sparse and pollinator service is chronically low.
• Annuals vs. Perennials: RA is generally more vital for annual plants, which must produce seeds before they die to ensure representation in the next generation. In contrast, long-lived perennials can withstand a poor pollination year and may avoid the costs of selfing to prioritize higher-quality outcrossed offspring in future years.
3. Evolutionary Trade-offs and Costs
While reproductive assurance provides a “safety net,” it comes with significant genetic and resource costs:
• Inbreeding Depression: The primary cost of RA is the reduced vigor and survival of selfed offspring due to the expression of deleterious recessive mutations. If the fitness cost of inbreeding is too high, RA may not provide a net benefit.
• Seed and Pollen Discounting: Using ovules or resources to produce selfed seeds might prevent a plant from producing higher-quality outcrossed seeds (seed discounting). Furthermore, selfing can reduce the amount of pollen available for export to other plants, decreasing the plant’s success as a “father” (pollen discounting).
• The Geitonogamy Constraint: Mating between different flowers on the same plant (geitonogamy) is genetically equivalent to selfing but typically does not provide reproductive assurance because it still requires a pollinator to move the pollen.
4. Impact of Environmental Disturbance
Human-induced changes, such as habitat fragmentation and climate change, can intensify pollen limitation, making reproductive assurance an increasingly necessary strategy for short-term survival. However, chronic reliance on RA can lead to the evolution of a “selfing syndrome”—where plants lose attractive traits like large petals and nectar—and may ultimately lead to a loss of genetic diversity, making the population more vulnerable to extinction in the long term.
9. Can plants recover lost genetic diversity?
Based on the sources, plants generally cannot easily recover lost genetic diversity, particularly once a lineage has transitioned to a strategy of predominant self-fertilization (selfing). This shift is widely characterized as an “evolutionary dead end” because it leads to a permanent reduction in genetic variation and adaptive potential.
The following factors explain the difficulty of recovering diversity and the rare circumstances under which it might be possible:
1. The Evolutionary Dead-End Hypothesis
The sources state that the transition from outcrossing to selfing is typically unidirectional and irreversible.
• Loss of Neutral Diversity: Self-fertilization reduces the effective population size (Ne)—often by half or more—which accelerates genetic drift and the loss of neutral genetic variants.
• Reduced Recombination: Selfing lineages have lower recombination rates, meaning they cannot easily create new genetic combinations or separate beneficial mutations from deleterious ones.
• Irreversibility: Once complex physiological mechanisms like self-incompatibility (SI) are lost, they are extremely unlikely to re-evolve, making a return to obligate outcrossing nearly impossible.
2. Recovery Through Gene Flow
While a single population may recover some diversity through gene flow—the influx of pollen or seeds from other populations—this is less effective in selfing species.
• Restricted Dispersal: Selfing species typically exhibit very low rates of pollen dispersal between populations, limiting the “rescue” effect of new genes from the outside.
• Metapopulation Dynamics: In systems where local populations frequently go extinct, recolonization can occasionally restore diversity, but selfing species often lack the genetic connectivity needed for robust recovery.
3. Purging vs. Mutation Accumulation
Self-fertilization does allow plants to purge deleterious recessive mutations by exposing them in a homozygous state.
• Fitness Maintenance: If a population persists long enough to purge its “genetic load,” it may evolve low inbreeding depression, which helps the population survive in the short term.
• Mutational Meltdown: However, this purging does not recover lost diversity; rather, the accumulation of mildly deleterious mutations and the loss of neutral variation may still lead to extinction over long evolutionary timescales.
4. Impact of Human Disturbance
Anthropogenic changes, such as habitat fragmentation and selective logging, make recovery even more difficult.
• Increased Inbreeding: Disturbance forces many plants toward selfing to ensure seed set (reproductive assurance), but this reduces the genetic diversity available for future adaptation.
• Isolation: By isolating populations, human disturbance cuts off the gene flow that would normally be the primary mechanism for a population to “recover” its genetic health.
In summary, while plants can occasionally offset the fitness costs of inbreeding by purging harmful mutations, the neutral genetic diversity and adaptive flexibility lost during the transition to selfing or through extreme population bottlenecks are rarely, if ever, recovered.
10. How do plants age and floral display size relate?
As plants age, they generally grow larger, which typically leads to an increase in the number of flowers produced during each reproductive episode. This positive correlation between plant age and floral display size has significant implications for reproductive success, pollinator behavior, and the evolution of plant mating systems.
The relationship between plant age and floral display involves the following key dynamics:
1. Mass Blooming and Geitonogamy
As plants grow older and produce larger floral displays (often referred to as mass blooming), the risk of geitonogamy—pollen transfer between different flowers on the same individual—increases.
• Pollinator Foraging: Pollinators are often attracted to larger displays because they offer increased floral rewards. However, once attracted, they tend to stay on the plant longer and visit multiple flowers in succession, which facilitates self-pollination.
• Mating Costs: Geitonogamy is generally maladaptive because it results in selfed offspring subject to inbreeding depression and leads to pollen discounting, where pollen is “wasted” on selfing instead of being exported to other plants.
2. Size-Dependent Gender Modification
Many plants exhibit size-dependent gender modification, where their sexual expression changes as they age and grow.
• Maternal Investment Costs: Producing seeds and fruits is energetically expensive. Consequently, many species develop as males when they are small and young, only transitioning to female function (producing seeds) once they reach a larger size capable of bearing those costs.
• Resource Allocation: In some species, such as Wurmbea dioica, larger plants are more likely to produce hermaphrodite flowers, while smaller individuals may remain purely male.
• Wind-Pollinated Exceptions: While animal-pollinated plants often become more “female” as they grow, some wind-pollinated species may become more male-biased with increased size. This is because taller, older plants can achieve greater fitness returns by dispersing pollen more effectively from a greater height.
3. Life-History Trade-offs (Annuals vs. Perennials)
The relationship between age and display size differs fundamentally between life forms:
• Perennials: These plants live for many years and grow progressively larger. Their large size makes them more prone to geitonogamy, which has likely acted as a selective force for the evolution of dioecy (separate sexes) or self-incompatibility to prevent the costs of mass-display selfing.
• Annuals: Short-lived plants must produce seeds before they die. They often have smaller displays and higher rates of autonomous selfing to ensure reproductive assurance, as they do not have the luxury of waiting for future reproductive seasons.
4. Clonal Architecture and Aging
In clonal plants, aging involves the expansion of the clonal colony. As the colony grows, the number of genetically identical flowering shoots (ramets) increases.
• Restricted Dispersal: As clones become larger, pollinators may move almost exclusively between genetically identical ramets, leading to extremely high rates of geitonogamy.
• Evolutionary Pressure: In species like Sagittaria latifolia, the mating costs associated with large clone sizes in stable habitats are thought to be a primary driver for the evolution of dioecy to eliminate the possibility of self-fertilization entirely.
In summary, as plants age and their floral displays grow, they face a “geitonogamy constraint” where the benefits of attracting more pollinators are increasingly offset by the genetic and paternal costs of self-pollination. This often forces older, larger plants to adopt more complex structural or sexual strategies to maintain outcrossing success.
11. Explain why separate sexes evolve in massive clones.
The evolution of separate sexes (dioecy) in massive clonal plants is primarily driven by the need to resolve the “geitonogamy constraint,” where large plant size or extensive clonal growth leads to high rates of self-fertilization and significant mating costs.
Separate sexes evolve in massive clones due to the following factors:
1. The Vulnerability to Geitonogamy
Massive clones consist of numerous genetically identical shoots (ramets) that often bloom simultaneously. Because pollinators typically visit multiple flowers or inflorescences on the same individual before moving to a new one, large clones experience high levels of geitonogamy (self-pollination between flowers on the same genotype). Unlike autonomous selfing within a single flower, geitonogamy is considered maladaptive because it requires a pollinator to occur but provides no reproductive assurance in the absence of mates.
2. High Maternal and Paternal Costs
The high incidence of geitonogamy in large clones imposes severe fitness penalties:
• Inbreeding Depression (Maternal Cost): Selfed seeds produced via geitonogamy often suffer from strong inbreeding depression, leading to reduced survival and vigor in offspring.
• Pollen Discounting (Paternal Cost): Pollen transported between flowers on the same clone is “wasted” and not available for export to other plants, significantly reducing the individual’s success as a male parent.
3. Release from the “Geitonogamy Constraint”
Separate sexes provide an ultimate evolutionary solution to these costs. By being exclusively male or female, a clone can grow to a massive size and produce large floral displays to maximize pollinator attraction without any risk of self-fertilization. This transition is often favored when the costs of inbreeding depression and pollen discounting in a hermaphroditic or monoecious state outweigh the advantages of having combined sexes.
4. Empirical Evidence: Sagittaria latifolia
The clonal aquatic plant Sagittaria latifolia serves as a key example of this transition:
• Habitat Correlation: Monoecious populations (hermaphroditic) typically occupy short-lived, disturbed habitats where clones remain small.
• Transition to Dioecy: In stable, competitive habitats where clones grow massive, the monoecious phenotype becomes highly maladaptive because over half the seeds produced can result from geitonogamy.
• Selective Advantage: In these massive clones, unisexual variants (males or females) spread because they avoid the high genetic load of selfed offspring, leading to the establishment of dioecious populations.
In summary, separate sexes evolve in massive clones as a strategic adaptation to decouple the benefits of large floral displays from the genetic costs of selfing, ensuring that all reproductive effort results in high-quality, outcrossed offspring.
12. What is synchronized protandry in vertical flower spikes?
Synchronized protandry is a specific form of dichogamy (temporal separation of sex functions) occurring in plants with vertical inflorescences, such as spikes or racemes, where the entire inflorescence acts as the reproductive unit.
This strategy relies on a precise coordination between flower maturation and pollinator behavior:
Mechanism of Synchronized Protandry
• Sequential Maturation: Individual flowers on the vertical spike mature sequentially from the bottom to the top.
• Sexual Phases: Because the flowers are protandrous, each flower first passes through a male phase (releasing pollen) before transitioning into a female phase (becoming receptive to pollen).
• Spatial Segregation: Due to the bottom-to-top maturation, the lower, older flowers on the spike are typically in their female phase, while the younger, upper flowers are in their male phase.
The Role of Pollinator Behavior
This strategy is highly effective because of the foraging patterns of pollinators, particularly bees:
• Upward Foraging: Bees generally begin foraging at the bottom of a vertical inflorescence and move upward.
• Efficient Pollen Transfer: By starting at the bottom, the bee first visits female-phase flowers, depositing “outcross” pollen it collected from a different plant. As it moves to the top of the spike, it visits male-phase flowers, picking up fresh pollen to carry to the next plant.
Evolutionary Benefits
Synchronized protandry evolved to resolve the conflict between a plant’s need for a large floral display and the risks of self-pollination:
• Reducing Geitonogamy: By segregating sex functions among flowers on the same plant, it minimizes geitonogamy—the transfer of pollen between different flowers on the same individual.
• Minimizing Sexual Interference: It reduces “sexual interference,” where a plant’s own male and female functions conflict, such as self-pollen “clogging” a stigma.
• Maximizing Paternal Fitness: It reduces pollen discounting, a cost where a plant’s own pollen is “wasted” on its own stigmas instead of being exported to other plants to sire seeds.
• Support for Large Displays: This mechanism allows plants to maintain larger, showier floral displays to attract more pollinators without paying the high genetic costs of inbreeding depression that would normally result from having many flowers open at once.
12. How do plants and animals differ in their avoidance of inbreeding?
Plants and animals differ fundamentally in their avoidance of inbreeding due to differences in mobility, modular growth, and sexual systems. While plants primarily utilize physiological and mechanical barriers within sedentary structures, animals rely heavily on behavioral adaptations and pre-mating movement.
According to the sources, the key differences in inbreeding avoidance include:
1. Primary Strategies of Avoidance
• Plants (Mechanical and Chemical): Because most plants are immobile and hermaphroditic, they face a high risk of “selfing”. They avoid this through Self-Incompatibility (SI), a molecular recognition system where the plant rejects its own pollen if it matches alleles at the S-locus. They also use floral design strategies like herkogamy (spatial separation of sex organs) and dichogamy (temporal separation of male and female functions) to prevent self-pollination.
• Animals (Movement and Discrimination): Most animals are gonochoric (separate sexes) and mobile, which naturally lowers the risk of inbreeding. Their primary mechanism is natal pre-mating dispersal, where juveniles move away from their place of birth before reaching reproductive maturity. Additionally, some animals use kin discrimination to identify and avoid mating with relatives.
2. Sexual Systems as Avoidance Mechanisms
• Separate Sexes: In plants, dioecy (separate male and female individuals) is a robust but relatively rare strategy (found in about 6% of species) evolved specifically to ensure outcrossing and avoid the genetic costs of selfing. In the animal kingdom, separate sexes are the standard, occurring in the vast majority of species, which fundamentally structures their mating systems toward outbreeding.
• Hermaphroditism: About 94% of flowering plants are cosexual (hermaphroditic), making inbreeding a constant potential outcome. In contrast, hermaphroditism is rare in animals (~5%–6%), occurring mostly in specific groups like snails or flatworms.
3. Post-Mating Avoidance
• Plants: After pollination, the “closed carpel” of angiosperms acts as a filter, allowing the plant to screen and reject unsuitable or self-pollen tubes during their growth through the style.
• Animals: Animals may utilize polyandry (female multiple mating), which enables sperm competition and cryptic female choice. These processes facilitate fertilization by unrelated or genetically compatible sperm even if a female has encountered a related mate.
4. Conceptual and Evolutionary Perspectives
• Inbreeding as “Adaptive” vs. “Negative”: In plant research, self-fertilization is often viewed as adaptive due to its automatic transmission advantage (Fisher’s 1.5-fold advantage) and its role in reproductive assurance when pollinators are scarce.
• Inbreeding as “Maladaptive”: In animal research, inbreeding is almost universally considered negative or a “maladaptive outcome” resulting from constraints like small population size. Animal species rarely show active inbreeding preference; instead, they show inbreeding tolerance only when mate encounter rates are very low.
Summary of Key Differences
| Feature | Plants | Animals |
|---|---|---|
| Main Barrier | Physiological (Self-Incompatibility) | Behavioral (Dispersal/Discrimination) |
| Timing | Post-pollination filtering | Pre-mating dispersal |
| Hermaphroditism | Overwhelmingly common (~94%) | Rare (~5.6%) |
| Selfing View | Often seen as an adaptive “safety net” | Generally seen as maladaptive |
| Spatial Driver | Baker’s Law (long-distance colonization) | Pre-mating dispersal (kin avoidance) |
13. Why do some plants still use mixed mating?
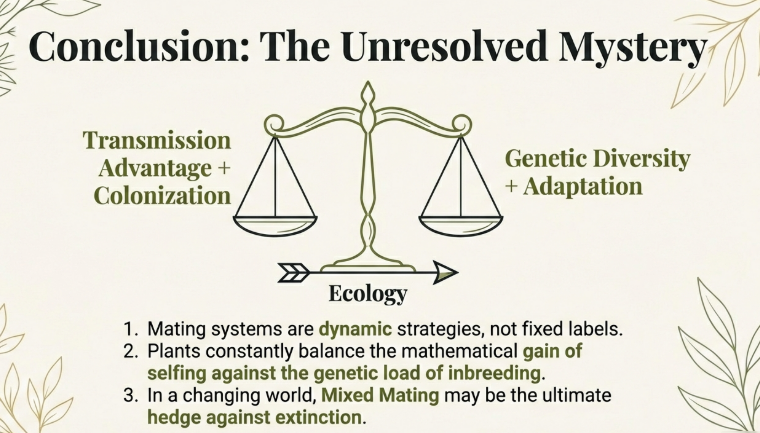
According to the sources, mixed mating—a system where populations produce offspring through both self-fertilization (selfing) and cross-fertilization (outcrossing)—is an “evolutionary enigma” because classical theoretical models predict that mating systems should instead evolve toward either extreme (predominant outcrossing or predominant selfing). Despite these predictions, approximately 42% of flowering plants exhibit a mixed mating system in nature.
Plants utilize mixed mating strategies for several adaptive and non-adaptive reasons:
1. Reproductive Assurance (The “Safety Net”)
The most significant adaptive reason for mixed mating is reproductive assurance. This strategy ensures that a plant can still produce seeds (fitness) even when pollination services are unreliable or unpredictable due to a scarcity of pollinators or mates.
• Delayed Selfing: Many plants employ “delayed selfing,” where mechanisms allow the flower to self-fertilize only after opportunities for outcrossing have passed. This is considered the “best of both worlds,” prioritizing high-quality outcrossed seeds but “failing safe” to selfed seeds if necessary.
• Baker’s Law: Mixed mating is a critical advantage for colonizing species or those at the margins of their range, as it allows a single individual to establish a new population without needing a mate.
2. Maladaptive Geitonogamy (The “Cost of Being Showy”)
In many cases, the selfing component of a mixed mating system is not an adaptive choice but a non-adaptive cost of having large floral displays designed to attract pollinators.
• Pollinator Foraging: When a plant produces many flowers to be more noticeable, pollinators often visit multiple flowers on that same individual. This leads to geitonogamy (pollen transfer between flowers on the same plant), which is genetically equivalent to selfing but provides no reproductive assurance because it still requires a pollinator to occur.
• Selection Balance: Selection against this maladaptive selfing is often hindered because the benefits of attracting more pollinators through large displays outweigh the genetic costs of the resulting inbreeding.
3. Evolutionary Trade-offs and Genetic Benefits
Mixed mating persists due to a complex balance of selective forces:
• Transmission Advantage: Selfing provides an automatic 1.5-fold gene transmission advantage; a selfing individual transmits more copies of its genes to the next generation than an outcrosser.
• Counteracting Costs: This advantage is balanced by inbreeding depression (reduced fitness of inbred offspring) and pollen discounting (selfing reduces the amount of pollen available to sire seeds on other plants). If inbreeding depression is intermediate (around 0.5), mixed mating can be maintained as a stable strategy.
4. Ecological and Environmental Stabilization
Fluctuating environments can prevent a population from settling on a single mating strategy:
• Environmental Stochasticity: Variations in pollinator abundance, plant density, and habitat stability can favor different mating strategies in different years or locations. In unstable environments, cleistogamy (producing both open and permanently closed, selfing flowers) may act as a bet-hedging strategy.
• Human Disturbance: Anthropogenic factors like habitat fragmentation and climate change can reduce mate or pollinator availability, forcing populations that were historically outcrossing to adopt mixed mating to survive.
5. Context-Dependent Mating
The outcrossing rate is often not a fixed trait of a species but is context-dependent, varying based on the local mating neighborhood. Factors such as the relative abundance of open flowers on a target plant compared to its neighbors, the specific identity of available pollinators, and the density of the population can all shift a plant’s mating behavior toward or away from selfing.
In summary, plants continue to use mixed mating because it offers a flexible balance between maximizing genetic transmission and offspring quality while providing a vital demographic insurance policy against the risks of an unpredictable environment.
14. Can selfing lead to a species’ extinction?
The transition to self-fertilization (selfing) is widely regarded as an “evolutionary dead end” that significantly increases a species’ risk of extinction over long evolutionary timescales. While selfing offers immediate short-term reproductive advantages, it triggers a cascade of genetic and demographic consequences that compromise long-term survival.
The Evolutionary “Dead-End” Hypothesis
The sources explain that selfing leads to extinction through several primary mechanisms:
• Loss of Genetic Diversity: Self-fertilization reduces the effective population size (Ne), often by half or more. This accelerates genetic drift, causing the rapid loss of neutral genetic variants and reducing the overall genetic diversity within a population.
• Reduced Recombination: Selfing lineages have much lower recombination rates than outcrossers. This limits the ability of a species to create new genetic combinations or to separate beneficial mutations from deleterious ones, which severely reduces adaptive potential—the capacity to evolve in response to changing environments or new threats.
• Mutational Meltdown: Because selection is less efficient in small, inbred populations, selfing species are prone to the accumulation and fixation of mildly deleterious mutations. This process can lead to a “mutational meltdown,” where the collective genetic load becomes so heavy that population fitness declines toward extinction.
The Paradox of Short-Term Success vs. Long-Term Failure
Selfing persists in nature because it is driven by individual selection that ignores the long-term survival of the species:
• Transmission Advantage: An allele for selfing has a 1.5-fold “automatic gene transmission advantage” (Fisher’s advantage), allowing it to spread rapidly through an outcrossing population even if it lowers “population fitness” or long-term survival.
• Reproductive Assurance: Selfing provides a vital “safety net” when pollinators or mates are scarce, ensuring seed production in the short term.
• Irreversibility: Once a lineage transitions to predominant selfing and loses complex outcrossing mechanisms (like self-incompatibility), the change is almost always unidirectional and irreversible.
Evidence from the Evolutionary Record
Phylogenetic data provide empirical support for the dead-end hypothesis:
• Lineage Deficit: Large-scale analyses of plant and animal (e.g., nematode) phylogenies show a significant deficit of “old” selfing lineages. This suggests that while lineages transition to selfing frequently, they go extinct at a much faster rate than their outcrossing counterparts.
• Sister Taxa Comparisons: Dioecious (separate sex) clades often contain fewer species than their outcrossing sister taxa, which some researchers suggest may be due to higher extinction risks associated with specialized mating systems.
Impact of Environmental Disturbance
The sources suggest that human environmental disturbance (e.g., habitat fragmentation, climate change) may accelerate this process by forcing more species toward selfing for immediate survival. While this provides short-term reproductive assurance, it may trap species in the “selfing syndrome,” leading to a loss of genetic flexibility and eventual extinction as the known deleterious costs of inbreeding are realized.
15. How does selfing affect a plant’s defenses against herbivores?
The transition from outcrossing to self-fertilization (selfing) significantly alters how plants deploy their defenses against herbivores. Research across the nightshade family (Solanaceae) indicates that selfing leads to a macroevolutionary shift from constitutive defenses (always present) to inducible defenses (activated only after an attack).
The impact of selfing on plant defense is characterized by the following shifts:
1. Shift from Constitutive to Inducible Defense
The repeated, unidirectional transition from ancestral self-incompatibility (obligate outcrossing) to self-compatibility (increased inbreeding) is strongly associated with the evolution of inducibility.
• Greater Inducibility: Resistance to herbivores is, on average, 63% more inducible in self-compatible (SC) taxa compared to self-incompatible (SI) taxa.
• Reduced Baseline Resistance: SC taxa tend to exhibit a moderate decrease in constitutive resistance compared to their outcrossing relatives. This means they invest less in “always-on” defenses and rely more on a plastic response after initial damage.
2. Evolutionary Trade-offs
The sources establish that constitutive and inducible strategies are evolutionary alternatives. There is a highly significant negative relationship between a species’ level of constitutive resistance and its inducibility.
• Plants that invest heavily in constitutive defenses save the risk of being unprotected but pay a constant resource cost.
• Selfing species appear to favor the inducible strategy, saving the cost of trait expression in the absence of herbivores but accepting the risk of initial damage.
3. Increased Specificity in Response
Contrary to the hypothesis that inbreeding might cause “developmental instability” or clumsy responses, self-compatible taxa actually show increased specificity in their induced resistance.
• Precision: SC taxa are better at evolving independent responses to different types of damage, such as distinguishing between mechanical wounding and actual herbivore consumption.
• Outcrossing Lack of Specificity: In contrast, SI (outcrossing) taxa show low specificity, with a strong correlation between their responses to mechanical wounding and real herbivory.
4. Ecological Drivers for the Defense Shift
The sources suggest that the relationship between selfing and inducible defense is an indirect result of the selective environments that accompany a shift to inbreeding:
• Marginal Habitats: Self-compatible taxa often persist in marginal habitats or range edges where pollinators and mates are scarce. These environments are often characterized by variable herbivory, which theoretical models predict will favor the evolution of phenotypic plasticity and inducible defenses.
• Resource Allocation: Induction of toxic metabolites in reward tissues (like nectar or pollen) could be disproportionately costly to obligately outcrossing plants that rely on pollinators. Selfing plants, being less dependent on these mutualists, may have more flexibility to deploy chemical defenses that might otherwise deter pollinators.
In summary, the transition to selfing drives a fundamental change in a plant’s “defensive syndrome,” moving away from permanent, high-level constitutive protection toward highly specific, plastic responses that are triggered only when an attack occurs.
16. How do pollinators influence the evolution of plant defense strategies?
Pollinators influence the evolution of plant defense strategies primarily by mediating the plant’s mating system, which in turn dictates how the plant allocates resources between permanent (constitutive) and reactive (inducible) defenses. Because pollinators and herbivores often interact with the same plant tissues—such as flowers and leaves—plants must evolve defensive syndromes that protect them from enemies without deterring their reproductive partners.
Pollinators influence plant defense evolution through the following mechanisms:
1. Indirect Influence via Mating System Transitions
The most significant influence identified in the sources is indirect: pollinator availability determines whether a plant remains an obligate outcrosser or evolves toward self-fertilization, which fundamentally shifts its defensive strategy.
• Shift from Constitutive to Inducible Defense: Research in the Solanaceae (nightshade) family shows that the loss of self-incompatibility (SI)—often caused by a scarcity of pollinators—drives a macroevolutionary shift from constitutive to inducible resistance. Self-compatible (SC) taxa exhibit resistance that is, on average, 63% more inducible than their outcrossing relatives.
• Reduced Baseline Investment: Because selfing plants often persist in marginal habitats or range edges where pollinators are unreliable, they may face more variable herbivory. This variability favors inducible defenses, which allow the plant to save the resource costs of defense expression when herbivores are absent.
2. Conflicting Selection on Floral Chemistry
Pollinators exert selective pressure on the chemical traits of a plant, which can conflict with the plant’s need to deter herbivores.
• Pollinator Deterrence: The induction of toxic or deterrent secondary metabolites in reward tissues, such as pollen and nectar, can be disproportionately costly to obligately outcrossing plants that depend entirely on pollinator visits.
• Defense Plasticity as a Solution: To resolve this conflict, plants may evolve highly specific inducible responses. Self-compatible taxa, being less dependent on these mutualists for every mating event, may have more flexibility to deploy chemical defenses that might otherwise deter pollinators.
3. Pollinator-Mediated Specificity
Pollinator-mediated selection appears to enhance the precision of a plant’s defensive response.
• Greater Precision in Selfers: Self-compatible taxa are better at evolving independent responses to different types of damage (e.g., distinguishing between mechanical wounding and actual herbivore consumption) compared to outcrossing taxa, which show lower specificity.
• Avoiding “Stigma Gunking”: In some cases, pollinators carry heterospecific pollen that can “clog” or “gunk” a stigma. The evolutionary pressure to maintain “clean” stigmas for successful mating may influence the structural and temporal deployment of floral defenses.
4. Specialized Antagonistic Mutualisms
In rare co-evolved systems, the pollinator and the herbivore are the same individual, leading to extreme defensive and reproductive adaptations.
• Obligate Antagonists: Examples include the yucca and yucca moth or figs and fig wasps. In these relationships, the moth is an obligate pollinator but its larvae are antagonistic herbivores that consume the plant’s seeds. This forces the plant to evolve strategies that balance the survival of its seeds with the necessity of the moth’s pollination services.
5. Influence of Pollination Mode
The method by which pollen is transferred also correlates with defensive structures.
• Biotic vs. Abiotic Pollination: Animal-pollinated flowers often have ovaries that are well-protected from the biting mouthparts of their pollinators, such as beetles.
• Wind Pollination and Dioecy: Wind-pollinated species, which do not need to attract pollinators, often evolve separate sexes (dioecy) and may invest differently in constitutive defenses because they lack the “pollinator deterrence” constraint found in animal-pollinated plants.
17. Why do self-compatible species show higher specificity in defense?
Self-compatible (SC) species show higher specificity in their defensive responses primarily because they have evolved to distinguish independently between different types of damage, such as mechanical wounding and actual herbivory. This allows them to “fine-tune” their defenses in an adaptive manner that is often absent in their self-incompatible (SI) relatives.
The following factors explain this increased specificity:
1. Evolutionary Independence of Responses
In self-incompatible (outcrossing) taxa, there is a strong and significant correlation between the plant’s response to mechanical wounding and its response to real herbivore damage. This indicates low specificity, as the plant essentially reacts the same way to any tissue loss. In contrast, in self-compatible taxa, these responses evolve independently, showing a non-significant correlation. This independence allows SC species to deploy highly tailored defenses specifically triggered by herbivore-associated cues rather than generic physical damage.
2. Rejection of the “Developmental Instability” Hypothesis
One might expect inbreeding in self-compatible species to cause “developmental instability”—clumsy or maladaptive plastic responses. However, the sources note that if SC plants were simply unstable, they would show lower specificity (uncontrolled, broad responses). The fact that they show greater specificity suggests that the shift to highly precise, inducible defenses is an adaptive evolutionary strategy rather than a negative byproduct of inbreeding.
3. Adaptation to Marginal Habitats
Self-compatible taxa are frequently found in marginal habitats or at species range edges where pollinators and mates are scarce. These environments are often characterized by variable and unpredictable herbivory. Theoretical models predict that such environments favor the evolution of highly specific phenotypic plasticity, allowing plants to save the costs of defense expression except when precisely needed.
4. Reduced Conflict with Pollinators
Inducible defenses can sometimes involve the production of toxic metabolites in reward tissues like nectar or pollen.
• For SI (Outcrossing) Taxa: Deploying these defenses is risky because they may deter the very pollinators the plant relies on for reproduction, which can be disproportionately costly.
• For SC (Inbreeding) Taxa: Because these plants are less dependent on pollinators for mating (relying instead on self-fertilization for reproductive assurance), they have more evolutionary flexibility to deploy specific and potentially deterrent chemical defenses.
In summary, the transition to self-compatibility allows plants to move away from broad, permanent (constitutive) protection toward highly specific, inducible syndromes that are precisely calibrated to the type of threat they encounter.
18. How do plants use sexual systems to prevent self-fertilization?
Plants use diverse sexual systems—the physical and functional distribution of male and female reproductive organs—to avoid the genetic and demographic costs of self-fertilization. While most plants are hermaphroditic, approximately 6% of species have evolved specialized sexual structures to ensure outcrossing.
Plants utilize the following sexual systems to prevent or limit selfing:
1. Dioecy (Separate Sexes)
Dioecy is the most effective sexual system for preventing self-fertilization because it involves populations composed of entirely separate male and female individuals.
• Obligate Outcrossing: In dioecious species, self-fertilization is morphologically impossible, making these plants obligately outcrossing.
• The Geitonogamy Ultimate Solution: Dioecy is often the evolutionary response to the “geitonogamy constraint”. In massive clones or large, showy plants, pollinators often move between many flowers on the same individual, causing high rates of maladaptive self-pollination. By being unisexual, a plant can produce a massive floral display to attract pollinators without any risk of siring its own seeds.
2. Monoecy (Separate Flowers on One Plant)
In monoecious systems, a single plant produces separate male and female flowers.
• Spatial Separation: This system spatially separates sex functions within the individual, which prevents intraflower self-pollination (selfing within the same flower).
• Limitations: While monoecy reduces some forms of selfing, it does not guarantee the prevention of self-fertilization. Pollen can still be transferred between different flowers on the same plant via wind or animal vectors, a process known as geitonogamy.
3. Gender Polymorphisms (Gynodioecy and Androdioecy)
Some plants utilize sexual systems where different individuals in a population have different “sex morphs” to promote outcrossing.
• Gynodioecy: This system consists of populations with unisexual females and cosexual (hermaphroditic) individuals. Females invade these populations because of an outbreeding advantage; they avoid the costs of inbreeding depression that the cosexual individuals might suffer from.
• Androdioecy: A very rare system in which populations are composed of cosexual and male plants. This system is often found in species with close dioecious relatives where females have evolved some ability to produce pollen.
4. Size-Dependent Gender Modification
Plants sometimes adjust their sexual system as they age or grow in size to manage the trade-offs of selfing.
• Cost Management: Because producing seeds and fruits is energetically expensive, many species develop as males when they are small and only transition to female function once they are large enough to bear the maternal costs.
• Wind-Pollination Bias: In some wind-pollinated monoecious species, plants may become more male-biased as they grow taller, as greater height allows for more effective pollen dispersal to other individuals, further reducing the chances of selfing within a dense patch.
5. Leaky Sex Expression
Even in systems designed to prevent selfing, such as dioecy, plants may exhibit “leaky” sex expression. This occurs when a unisexual plant (typically a female) produces flowers of the opposite sex. While this allows for self-fertilization, it is often seen as a reproductive assurance mechanism, ensuring that a plant can still produce seeds if no mates are available in a new or disturbed environment.
In summary, plants utilize these sexual systems to decouple the benefits of pollinator attraction from the genetic costs of inbreeding, ensuring that their reproductive effort results in high-quality, genetically diverse offspring.
19. Why is selfing called an evolutionary dead end?
Self-fertilization (selfing) is famously characterized as an “evolutionary dead end” because, while it provides immediate short-term reproductive advantages, it triggers genetic and demographic processes that significantly increase a lineage’s long-term risk of extinction. This hypothesis, originally proposed by Stebbins in 1957, suggests that selfing species are less able to adapt to environmental changes and are prone to a slow genetic deterioration.
Selfing is considered a dead end for the following reasons:
1. Loss of Genetic Diversity and Adaptive Potential
The most critical factor in the dead-end hypothesis is the permanent reduction of genetic variation.
• Reduced Effective Population Size (Ne): Inbreeding effectively halves the effective population size compared to an outcrossing population of the same census size. A smaller Ne accelerates genetic drift, leading to the rapid loss of neutral genetic variants and overall diversity.
• Limited Recombination: Selfing lineages have much lower rates of recombination than outcrossers. This prevents the creation of new genetic combinations and the separation of beneficial mutations from deleterious ones, which severely restricts the species’ ability to adapt to new threats or environmental shifts.
2. Accumulation of Deleterious Mutations
Selfing makes natural selection less efficient at maintaining the “health” of the genome.
• Weakened Selection: Because of reduced Ne and recombination, selection is less effective at eliminating disadvantageous mutations.
• Mutational Meltdown: Over time, inbreeding populations are predicted to slowly deteriorate by fixing mildly deleterious mutations. This accumulation can lead to a “mutational meltdown” where population fitness declines until the species can no longer persist.
3. Evolutionary Irreversibility
The transition from outcrossing to selfing is widely viewed as unidirectional.
• Loss of Complex Traits: Once a lineage loses the complex physiological and morphological mechanisms required for obligate outcrossing—such as self-incompatibility (SI) or separate sexes—these traits are extremely unlikely to re-evolve.
• Purging Constraints: If a population persists long enough to purge its “genetic load” of major deleterious mutations, it may evolve low inbreeding depression, which removes the primary evolutionary incentive to return to an outcrossing strategy.
4. Evidence from the Phylogenetic Record
Phylogenetic data provide empirical support for this theory by revealing a deficit of “old” selfing lineages.
• Short-Lived Lineages: Large-scale analyses of plant phylogenies (such as in the Solanaceae and Fabaceae families) and animal phylogenies (such as Caenorhabditis nematodes) show that selfing lineages tend to appear at the tips of the trees but rarely persist long enough to diversify significantly.
• Higher Extinction Rates: This pattern suggests that while lineages transition to selfing frequently, they go extinct at a much faster rate than their outcrossing counterparts.
In summary, selfing is an “evolutionary trap”: it is driven by individual selection for immediate gene transmission and reproductive assurance, but it ultimately robs the species of the genetic flexibility required for long-term survival in a changing world.
20. How do selfing species avoid mutational meltdown?
Self-fertilizing (selfing) species avoid mutational meltdown primarily through a process known as purging, where the increased homozygosity caused by inbreeding exposes deleterious recessive mutations to the forces of natural selection. However, the sources also indicate that this avoidance is often incomplete, leading to a long-term evolutionary “dead end” where many selfing lineages eventually go extinct.
Selfing species manage their mutation load through the following mechanisms:
1. Purging the Genetic Load
The most effective way selfing species avoid mutational meltdown is by purging.
• Expression of Recessive Alleles: In outcrossing populations, harmful recessive mutations are “masked” in a heterozygous state. Selfing rapidly increases the frequency of homozygotes, forcing these recessive mutations to be expressed in the phenotype.
• Selective Removal: Because these mutations often reduce survival or fertility (inbreeding depression), the individuals carrying them are less likely to reproduce. This allows natural selection to identify and eliminate these harmful alleles from the population’s gene pool.
2. The Rate of Inbreeding (“Slow” vs. “Fast”)
The efficiency of purging depends heavily on the rate at which inbreeding occurs:
• “Slow” Inbreeding: Matings between relatives (such as half-sibs) allow for more efficient purging. Because the effective population size (Ne) remains relatively higher than in extreme selfing, natural selection is more precise in removing disadvantageous alleles while preserving advantageous ones.
• “Fast” Inbreeding: Extreme or obligate selfing exposes mutations very quickly but also drastically reduces Ne (often by half or more). This strengthens the power of genetic drift, which can accidentally “fix” mildly deleterious mutations within a population faster than selection can remove them, potentially triggering a mutational meltdown.
3. Survival through Low Inbreeding Depression
If a selfing population persists long enough to successfully purge its genetic load, it may evolve low inbreeding depression. Once the most harmful mutations are gone, the primary genetic cost of selfing is removed, allowing the population to maintain high fitness in the short term despite its lack of genetic diversity.
4. The “Dead-End” Constraint
While purging helps avoid immediate fitness collapse, it does not fully protect a species from long-term extinction:
• Loss of Adaptive Potential: While purging removes deleterious variants, the combination of selfing and genetic drift also leads to a rapid loss of neutral genetic diversity.
• Reduced Recombination: Selfing lineages have significantly lower recombination rates. This prevents them from “shuffling” their genes to create new adaptive combinations or from separating beneficial mutations from mildly harmful ones.
• Macroevolutionary Evidence: Phylogenetic data show a deficit of old selfing lineages, suggesting that while these species can survive for some time by purging their load, they eventually succumb to extinction because they cannot adapt to changing environmental conditions or new threats.
In summary, selfing species avoid mutational meltdown in the short term by using homozygosity to filter out harmful mutations through purging. However, the sources emphasize that this strategy is often a temporary survival mechanism that eventually leads to an evolutionary dead end due to the permanent loss of genetic flexibility.
21. Why do large floral displays sometimes reduce a plant’s paternal fitness?
Large floral displays, while evolved to maximize pollinator attraction, can sometimes reduce a plant’s paternal fitness (its success in siring seeds on other plants) due to a phenomenon known as pollen discounting. This occurs primarily through increased rates of geitonogamy—the transfer of pollen between different flowers on the same individual plant.
Large floral displays reduce paternal fitness through the following mechanisms:
1. Increased Geitonogamy
Large floral displays offer increased visual signals and rewards, which attract more pollinators. However, once a pollinator is attracted to a large display, it tends to remain on that individual for a longer duration, visiting multiple flowers in succession. This foraging behavior facilitates the transfer of pollen between flowers on the same plant.
2. Pollen Discounting
The primary paternal cost of geitonogamy is pollen discounting, which is the reduction in an individual’s outcrossed siring success caused by its own self-pollination.
• Wasted Pollen: Pollen that is transported between flowers on the same plant is “wasted” because it is no longer available to be exported to other individuals in the population.
• Reduced Male Fertility: Because the amount of pollen a plant produces is finite, every grain used in geitonogamous selfing represents a lost mating opportunity to sire higher-quality, outcrossed seeds elsewhere.
3. Lack of Reproductive Assurance
Unlike autonomous self-pollination within a single flower (which ensures seed set when pollinators are scarce), geitonogamy requires a pollinator to occur. Consequently, it is often viewed as a maladaptive consequence of being showy; the plant pays the paternal cost of reduced outcrossing success without receiving the demographic “safety net” that other forms of selfing provide.
4. Evolutionary Solutions to the Conflict
Because the “geitonogamy constraint” creates a conflict between attraction and mating quality, plants have evolved several strategies to decouple the benefits of large displays from the paternal costs of selfing:
• Synchronized Protandry: In vertical inflorescences, plants often mature flowers from the bottom up and time them so that a pollinator picks up pollen from male-phase flowers at the top after it has already deposited outcross pollen on female-phase flowers at the bottom.
• Dioecy (Separate Sexes): In massive clones where geitonogamy would be inevitable and highly damaging, species may evolve separate male and female individuals, allowing for massive floral displays with zero risk of self-pollination.
• Structural Barriers: Mechanisms like herkogamy (spatial separation of sex organs) and dichogamy (temporal separation) function not just to prevent inbreeding, but to promote more proficient pollen dispersal and minimize sexual interference.
22. How do plants shift from constitutive to inducible defense?
Plants shift from constitutive to inducible defense strategies primarily as a result of macroevolutionary transitions in their mating systems, specifically the move from obligate outcrossing (self-incompatibility) to increased inbreeding (self-compatibility).
This shift involves the following mechanisms and ecological drivers:
1. The Mating System Transition
Research across the Solanaceae (nightshade) family demonstrates that the repeated loss of self-incompatibility (SI) leads to a fundamental change in how plants deploy resistance traits.
• Constitutive Defense: These are “always-on” defenses, such as physical toughness or constant toxin production. These are more prevalent in self-incompatible (outcrossing) taxa.
• Inducible Defense: These are reactive defenses triggered only after an initial herbivore attack, saving the plant the metabolic cost of defense expression when enemies are absent. Self-compatible (SC) taxa exhibit resistance that is, on average, 63% more inducible than their outcrossing relatives.
2. Ecological Drivers of Inducibility
The shift is often an indirect result of the selective environments that accompany a move toward selfing:
• Marginal Habitats and Variable Attack: Selfing species frequently persist in marginal habitats or at species range edges where both mates and pollinators are scarce. These environments often feature variable and unpredictable herbivory. Theoretical models predict that such variability favors phenotypic plasticity, allowing plants to remain undefended until the risk of attack is confirmed.
• Resource Allocation and Trade-offs: There is a strong macroevolutionary trade-off between the two strategies; as a species evolves higher inducibility, it typically shows a moderate decrease in its baseline constitutive resistance.
3. Resolution of Pollinator Conflicts
Inducible defenses provide a solution to the conflict between attracting pollinators and deterring herbivores:
• Pollinator Deterrence: Many chemical defenses are toxic or deterrent to pollinators if they are expressed in reward tissues like nectar or pollen.
• Flexibility for Selfers: Obligately outcrossing plants pay a high cost if they deter pollinators. Because SC plants are less dependent on pollinators for reproduction (due to reproductive assurance through selfing), they have more evolutionary flexibility to deploy chemical defenses that might otherwise be too costly for outcrossers to maintain.
4. Increased Specificity of Response
The transition to a more inducible strategy is also associated with a move toward higher specificity.
• Precise Recognition: Self-compatible taxa have evolved the ability to independently distinguish between mechanical wounding (e.g., wind damage) and actual herbivore consumption.
• Low Specificity in Outcrossers: In contrast, self-incompatible (outcrossing) taxa show a lack of independence in these responses, reacting similarly to any form of tissue loss.
In summary, the transition from outcrossing to selfing allows plants to move away from permanent, high-level protection and toward highly specific, plastic responses that are precisely calibrated to the threats they encounter in unpredictable environments.
23. Do animal-pollinated plants have higher inducibility than wind-pollinated ones?
Animal-pollinated plants do not necessarily have higher inducibility than wind-pollinated ones. In fact, for many animal-pollinated species, the need to attract pollinators acts as a constraint that limits the evolution of inducible defenses.
The sources provide the following insights into how pollination and mating systems influence defense strategies:
1. The “Pollinator Deterrence” Constraint
In animal-pollinated species, there is often a conflict between deterring herbivores and attracting pollinators.
• Costly Induction: For obligately outcrossing plants (which rely entirely on pollinators), the induction of toxic or deterrent metabolites in reward tissues like nectar or pollen can be disproportionately costly because it may drive away the very pollinators needed for reproduction.
• Lower Inducibility in Outcrossers: Because of this constraint, self-incompatible (obligately outcrossing) animal-pollinated taxa tend to exhibit lower inducibility and rely more on constitutive (permanent) defenses.
2. Selfing and Higher Inducibility
The sources highlight that the transition to self-fertilization—which often occurs in animal-pollinated lineages when pollinators or mates become scarce—drives a shift toward inducibility.
• 63% Increase: Research in the nightshade family (Solanaceae) shows that plant resistance to herbivores is, on average, 63% more inducible in self-compatible (SC) taxa compared to their self-incompatible (SI) outcrossing relatives.
• Flexibility: Because selfing plants are less dependent on pollinators for every mating event, they have more evolutionary flexibility to deploy chemical defenses that might otherwise deter mutualists.
3. Wind-Pollinated Species
The sources suggest that wind-pollinated plants occupy a different defensive niche because they are entirely free from the need to manage pollinator behavior.
• Lack of Deterrence Constraint: Wind-pollinated species do not have to worry about “pollinator deterrence”.
• Defense Investment: While the sources do not provide a direct statistical comparison of inducibility between animal and wind pollination across all families, they note that wind-pollinated species may invest differently in constitutive defenses because they lack the specific mechanical and chemical constraints imposed by animal vectors.
Summary Table of Defense Strategies
| Pollination/Mating Type | Pollinator Dependency | Primary Defense Strategy | Inducibility Level |
|---|---|---|---|
| Outcrossing Animal-Pollinated (SI) | High | Constitutive (to avoid deterring pollinators) | Lower |
| Selfing Animal-Pollinated (SC) | Low | Inducible (for flexibility in marginal habitats) | Higher (63% more) |
| Wind-Pollinated | None | Varied (often Constitutive) | High Flexibility (No deterrence cost) |
In conclusion, obligately animal-pollinated plants actually have lower inducibility than selfing species, and wind-pollinated plants lack the primary evolutionary barrier that keeps inducibility low in many animal-pollinated populations.
24. How do wind-pollinated flowers differ structurally from animal-pollinated ones?
Wind-pollinated flowers and animal-pollinated flowers differ significantly in their structural adaptations, as their designs are shaped by the specific requirements of their pollen vectors—either abiotic (wind) or biotic (animals like bees, birds, and bats).
The key structural differences include:
1. Attractive Structures and Rewards
• Animal-Pollinated: These flowers function as “attraction strategies”. They possess large, showy petals (or tepals) often characterized by bright colors, ultraviolet nectar guides, and complex shapes to attract and position pollinators. They also produce scents (sweet, musty, or foul) and rewards such as nectar and edible pollen.
• Wind-Pollinated: These flowers are generally small, inconspicuous, and green. They typically lack petals and sepals (the perianth) and do not produce scent or nectar because they do not need to attract animal vectors.
2. Pollen Production and Characteristics
• Animal-Pollinated: These plants produce relatively smaller amounts of pollen because the transfer is more targeted. In some specialized cases, like orchids, pollen is packaged into clusters called pollinia that stick to the insect.
• Wind-Pollinated: Because wind dispersal is non-directional and random, these plants produce enormous numbers of relatively small pollen grains to increase the probability of reaching a receptive stigma.
3. Stigma and Anther Anatomy
• Animal-Pollinated: The sex organs are precisely positioned (often using herkogamy, or spatial separation) to ensure they contact specific parts of a pollinator’s body. Some may have protected ovaries to prevent damage from the biting mouthparts of pollinators like beetles.
• Wind-Pollinated: The stigmas are often large and feathery, providing a greater surface area to “catch” drifting pollen grains from the air. Additionally, some species evolve elongated peduncles (flower stalks) to elevate male flowers, promoting more effective wind dispersal.
4. Sexual Systems and Plant Architecture
• Animal-Pollinated: The vast majority of these species are hermaphroditic (cosexual), as it is efficient to both donate and receive pollen during a single pollinator visit.
• Wind-Pollinated: These species are more frequently dioecious (separate male and female individuals) or monoecious (separate male and female flowers on one plant). This separation helps prevent “pollen clogging,” where a plant’s own pollen lands on its stigmas and interferes with outcrossing.
5. Phenology (Timing)
• Wind-Pollinated: These plants often bloom early in the growing season, frequently before their leaves have fully emerged, to ensure that foliage does not interfere with the movement of pollen through the air.
• Animal-Pollinated: Their flowering is timed to coincide with the activity periods of their specific animal vectors.
25. What is the evolutionary advantage of vegetative reproduction and apomixis?
Asexual reproduction in plants occurs in two fundamental forms: vegetative reproduction and agamospermy (apomixis). Both strategies provide significant evolutionary advantages related to genetic transmission efficiency, demographic survival, and reproductive assurance.
The primary advantages include:
1. Genetic Transmission Advantage
The most significant genetic benefit of asexual reproduction is the twofold transmission advantage. While an outcrossing sexual plant contributes only half of its genes to each offspring, an asexual plant transmits its entire genome to its progeny. This allows an asexual mutant or lineage to spread rapidly within a population because it is twice as efficient at propagating its genes.
2. Perennialization and Survival (Vegetative Reproduction)
Vegetative reproduction, common in perennial plants, acts as a mechanism for perennialize and expand.
• Seasonal Persistence: It allows plants to survive and persist from one growing season to the next in a specific location.
• Biomass Expansion: Plants can expand their size and cover wide areas quickly through structural modifications like rhizomes, stolons, or runners.
• Clonal Colonies: This process gives rise to clonal colonies where genetically identical individuals (ramets) occupy a habitat, increasing the likelihood of the genotype’s long-term survival.
3. Reproductive Assurance and Colonization
Asexual reproduction provides a vital demographic “safety net” known as reproductive assurance.
• Independent Reproduction: Apomixis allows for the formation and dispersal of seeds without fertilization. This ensures that a plant can still produce offspring even when pollinators are absent or potential mates are scarce.
• Baker’s Law: These strategies are critical for colonizing new habitats. Consistent with Baker’s Law, a single individual capable of asexual reproduction can establish a new population after long-distance dispersal without needing a mate or a specific animal vector.
4. Preservation of “Genetic Gold”
Because asexual offspring are genetically identical to their parent, these strategies preserve successful genotypes. In sexual reproduction, meiosis and fertilization rearrange genes, which can break up advantageous combinations of traits. Apomixis has been described as “genetic gold” because it allows a plant to replicate a highly fit genotype perfectly without the risk of genetic dilution or “scrambling”.
5. Resource Efficiency
Asexual strategies can be more resource-efficient than sexual reproduction. For example, apomictic seeds can be dispersed over long distances like sexual seeds but avoid the “cost of sex”. Vegetative reproduction allows for survival and expansion without the high energetic costs associated with producing complex floral displays, nectar, or large amounts of pollen to attract animal pollinators.
26. How do pollination syndromes like ‘bee’ or ‘moth’ differ?
Pollination syndromes are suites of related floral traits—such as shape, size, color, odor, and reward type—that have evolved through pollinator-mediated selection to attract specific classes of pollen vectors. While both bees and moths are important insect pollinators, their respective syndromes differ significantly based on their sensory capabilities, foraging behaviors, and metabolic needs.
The primary differences between the bee and moth syndromes include:
1. Color and Visual Cues
• Bee Syndrome: These flowers typically feature yellow or blue colors. A critical distinction is the presence of ultraviolet (UV) nectar guides, which are visible to bees but invisible to humans, helping to direct the insect to the center of the flower.
• Moth Syndrome: Because most moth species are nocturnal or crepuscular (active at twilight), their flowers tend to be white or light-colored to remain visible in low-light conditions. They generally lack the complex UV guides found in bee-pollinated species.
2. Scent and Timing
• Bee Syndrome: Bee flowers are generally diurnal (open during the day) and often produce a scent to attract foragers.
• Moth Syndrome: These flowers are primarily nocturnal or crepuscular, opening in the evening or night. They emit a strong, sweet scent specifically during the evening or night to attract moths over long distances in the dark.
3. Floral Shape and Morphology
• Bee Syndrome: These flowers are designed to accommodate landing. They often have sturdy structures or landing platforms where bees can perch while foraging.
• Moth Syndrome: Many moth-pollinated flowers, such as those visited by hawk moths (Sphingidae), have tubular corollas. Hawk moths forage similarly to hummingbirds, hovering in front of flowers with rapid wingbeats rather than landing, which allows them to reach nectar hidden in narrow tubes or spurs with their long proboscis.
4. Rewards (Nectar and Pollen)
• Bee Syndrome: Plants offer nectar, pollen, or both as rewards. The sugar in the nectar of bee-pollinated flowers tends to be sucrose-dominated. Some specialized “oil-collecting” bees visit flowers with special glands that provide oils instead of or in addition to nectar.
• Moth Syndrome: These flowers produce copious amounts of nectar. This is necessary to provide the high energy required to power the rapid wingbeats and high metabolic rates needed for hovering flight.
Summary of Differences
| Feature | Bee Syndrome | Moth Syndrome |
|---|---|---|
| Primary Colors | Yellow, blue, purple | White or pale |
| Visual Guides | Ultraviolet (UV) guides | Generally absent |
| Scent | Often present; diurnal | Strong, sweet; nocturnal |
| Timing | Day-opening | Night-opening |
| Flower Shape | Often sturdy with landing platform | Tubular corollas; often pendulous |
| Foraging Style | Landing and crawling | Hovering (hawk moths) |
| Rewards | Nectar (sucrose) and pollen | Large volumes of nectar |
In summary, the bee syndrome emphasizes daytime visibility and structural support for landing insects, while the moth syndrome focuses on nocturnal attraction through scent and pale colors, providing the high-volume nectar rewards necessary for the energy-intensive hovering flight of many moth species.
27. How does human-induced habitat fragmentation trigger more selfing?
Human-induced habitat fragmentation triggers increased rates of self-fertilization (selfing) primarily by disrupting plant-pollinator interactions and reducing the availability of potential mates. According to the sources, these anthropogenic changes create a cascade of ecological and evolutionary shifts that move plants away from outcrossing strategies:
1. Disruption of Pollination Services
Habitat fragmentation often leads to a reduction in pollinator abundance and diversity. When pollinators are scarce, the amount of outcross pollen arriving on a plant’s stigma decreases, which is known as outcross pollen limitation. In self-compatible species, this lack of cross-pollen allows self-pollen to fertilize a higher proportion of ovules by default.
2. Reduced Mate Availability
Fragmentation breaks large, contiguous plant populations into smaller, sparser, and more isolated patches.
• Fewer Mates: Small populations contain fewer unrelated conspecific individuals, limiting the number of potential outcross parents.
• Decreased Attraction: Sparser plant populations are often less noticeable to pollinators, further reducing the frequency of floral visits that would normally facilitate cross-pollination.
3. Selection for Reproductive Assurance (RA)
When human disturbance causes chronic outcross pollen limitation, it generates a strong selective pressure for reproductive assurance.
• The Safety Net: Selfing acts as a demographic “safety net,” ensuring the plant can still produce seeds even when mates or pollinators are unreliable or absent.
• Prior Selfing: In severely fragmented habitats where both pollinators and mates are consistently scarce, the sources suggest that prior selfing—where a flower self-pollinates before it even opens—is favored because it provides high RA and saves the metabolic costs of maintaining long-lived, attractive flowers.
4. Changes in Plant Architecture and Foraging
Fragmentation can alter plant size and floral display, which indirectly influences mating. For example, in fragmented urban populations of Crepis sancta, plants were found to be larger with showier displays due to reduced competition, which led to higher rates of geitonogamy (selfing between different flowers on the same plant) as pollinators spent more time foraging on a single individual.
5. Evolutionary Consequences
Over time, chronic fragmentation can lead to the evolution of the “selfing syndrome,” characterized by a reduction in attractive floral traits like petal size and nectar production. While this shift provides short-term survival through reproductive assurance, the sources caution it can lead to a permanent loss of genetic diversity and adaptive potential, potentially making the species an “evolutionary dead end” prone to extinction.
Empirical meta-analysis of 22 studies shows that outcrossing rates are significantly lower in human-disturbed habitats across various species and life forms, particularly in trees and woody shrubs.
28. What are the common methods of asexual reproduction in plants?
Asexual reproduction in plants occurs when offspring are produced by a single parent without the fusion of male and female gametes, resulting in new individuals that are genetically identical to the parent. Asexual reproduction occurs in two fundamental forms: vegetative reproduction and agamospermy (apomixis).
1. Vegetative Reproduction
This process is most common in perennial plants and utilizes structural modifications of the stem, roots, or leaves to perennialize the plant and expand its size. This leads to the formation of a clonal colony where genetically identical individuals, known as ramets, persist in a location.
Common natural structures and methods include:
• Rhizomes: Modified underground stems that can separate at the growing tips to form new plants (e.g., iris, polypody, ginger).
• Runners and Stolons: Prostrate aerial stems that produce new ramets at their nodes, often covering wide areas quickly (e.g., strawberries, many grasses).
• Bulbs, Tubers, and Corms: Plants like onions and hyacinths divide underground bulbs; potatoes reproduce through tubers; and gladioli use corms.
• Adventitious Buds and Suckering: Buds can form on roots or damaged stems (like tree stumps) and develop into new shoots. Suckering is the regeneration of a plant by shoots arising from an existing root system (e.g., elm, roses).
• Gemmae: Single cells or masses of cells that detach from the parent to form new clonal individuals, common in liverworts, mosses, and some higher plants like Drosera.
• Fragmentation and Regeneration: New individuals can grow from separated pieces of the original plant.
2. Agamospermy (Apomixis)
This form of reproduction is a replacement for sexual reproduction and involves the formation and dispersal of seeds without fertilization.
• Asexual Seeds: Species such as dandelions, hawkweeds, and Kentucky blue grass produce seeds that do not originate from the fertilization of embryos.
• Pseudogamy: A specific form of apomixis where pollination is required to initiate embryo growth, but the pollen contributes no genetic material to the offspring.
• Bulbils and Plantlets: Some plants produce bulbils instead of flowers or generate a plantlet in replacement of a seed to produce new cloned individuals.
3. Horticultural (Human-Enhanced) Methods
Humans often enhance natural processes to propagate cultivars with desirable characteristics that might not “come true” from seeds. These common methods include:
• Cuttings: Rooting a branch cut from a mother plant.
• Grafting: Joining a stem or bud from one plant onto the rooted stem of another.
• Layering and Division: Techniques to separate and root parts of a plant while they are still attached or recently divided.
• Laboratory Tissue Cloning: Artificial propagation using tissue cultures to create many identical clones in a controlled environment.
While asexual reproduction is highly efficient for gene transmission and colonization, the sources note a major disadvantage is the transmission of pathogens from parent to offspring, a risk that is less common in seed-based sexual reproduction.
29. How do butterflies and moths differ as pollinators for plants?
Butterflies and moths, while both members of the order Lepidoptera, have evolved distinct pollination syndromes with different floral adaptations, timing, and foraging behaviors.
Following are the primary differences between these two groups as pollinators:
1. Timing and Visual Cues
• Butterflies (Diurnal): Butterflies are active during the day, and the flowers they pollinate reflect this. These flowers tend to be large, showy, and brightly colored, frequently in shades of pink or lavender. They often possess simple nectar guides to direct the butterfly to the reward.
• Moths (Nocturnal/Crepuscular): Most pollinating moths, such as hawk moths (Sphingidae), are active at night or during twilight. Consequently, moth-pollinated flowers are typically white and night-opening to be visible in low light.
2. Foraging and Structural Support
• Landing Platforms (Butterflies): Because butterflies typically need to land to feed, the flowers they visit frequently provide a landing area or platform.
• Hovering Behavior (Hawk Moths): Many important moth pollinators, particularly hawk moths, forage similarly to hummingbirds by hovering in front of flowers with rapid wingbeats rather than landing. This behavior is supported by flowers with tubular corollas.
3. Scent and Rewards
• Butterflies: These flowers are usually scented and offer primarily nectar as a reward, as butterflies generally do not digest pollen. Their long tongues allow them to reach nectaries hidden deep within narrow tubes or spurs.
• Moths: Moth flowers emit a strong, sweet scent specifically in the evening, night, or early morning to attract nocturnal foragers. Because hovering flight is metabolically expensive, these flowers produce copious amounts of nectar to fuel the moth’s high metabolic rates.
4. Impact on Plant Genetic Structure
• Short-Distance Flight: Lepidoptera (both butterflies and moths) generally tend to fly short distances between plants.
• Genetic Differentiation: Because of this restricted movement, plants pollinated by butterflies and moths often exhibit higher genetic differentiation (FST) among populations compared to plants pollinated by long-distance fliers like certain bees or birds.
5. Co-Evolutionary Traits
Both groups have participated in striking examples of co-evolution with plants. For instance, the yucca and yucca moth have a specialized relationship where the moth is both the obligate pollinator and an antagonistic herbivore. More generally, the development of long floral corollas in many species is a direct evolutionary response to the long proboscii (tongues) of butterflies and moths.
30. What is the two-fold transmission advantage of asexual reproduction?
The two-fold transmission advantage of asexual reproduction (also known as the “two-fold cost of sex”) refers to the genetic efficiency with which an asexual parent propagates its genome compared to an outcrossing sexual parent.
This advantage is defined by the following principles:
• Genetic Contribution per Offspring: In sexual reproduction (outcrossing), an individual contributes only 50% of its genes to each offspring, with the remaining 50% coming from the mating partner. In contrast, an asexual parent transmits 100% of its genome to its progeny, as the offspring are genetically identical to the parent.
• Two-Fold Efficiency: Because an asexual plant contributes twice as much genetic material to each individual offspring as an outcrosser, it is twice as efficient at propagating its genes. This mathematical edge allows an asexual lineage or mutant to spread rapidly through a population.
• Preservation of “Genetic Gold”: Asexual reproduction, particularly apomixis (asexual seed production), is described in the sources as “genetic gold”. This is because it replicates a highly fit genotype perfectly, preserving successful combinations of traits without the genetic “scrambling” that occurs during meiosis and fertilization in sexual reproduction.
• Contrast with Self-Fertilization: While self-fertilization (selfing) also provides a transmission advantage (Fisher’s advantage), it is typically only a 1.5-fold (50%) advantage rather than a two-fold one. This is because a selfing individual transmits two gametes to its own seeds but may also contribute gametes to other plants through its pollen.
In summary, the two-fold advantage represents the immediate evolutionary incentive for asexual reproduction, allowing a genotype to bypass the genetic dilution inherent in mating with another individual.
31. How do pollinators distinguish between bee and moth syndromes?
Pollinators distinguish between bee and moth syndromes by responding to distinct “suites of related traits”—including color, scent, timing, and structure—that have evolved through pollinator-mediated selection to match their specific sensory systems and foraging behaviors.
The primary ways pollinators distinguish between these two syndromes are:
1. Visual Cues: Color and UV Guides
• Bee Syndrome: Bees are attracted to flowers that are typically yellow or blue. A critical distinguishing feature is the presence of ultraviolet (UV) nectar guides, which are visible to bees but often invisible to other pollinators (and humans), helping to direct the bee to the center of the flower.
• Moth Syndrome: Since most pollinating moths are nocturnal or crepuscular (active at twilight), their flowers are generally white or light-colored to maximize visibility in low-light conditions. They are often large and showy but lack the complex UV patterns found in bee flowers.
2. Olfactory Cues: Scent
• Bee Syndrome: These flowers possess a scent to attract foragers during their active hours.
• Moth Syndrome: Moths are highly sensitive to olfactory signals. Moth-pollinated flowers emit a strong, sweet scent specifically during the evening, night, or early morning to attract nocturnal foragers over long distances.
3. Temporal Cues: Timing of Opening
• Bee Syndrome: These flowers are generally active during the day to coincide with bee foraging patterns.
• Moth Syndrome: These are characteristically night-opening flowers, remaining closed or less attractive during the day and becoming receptive when moths are active.
4. Morphological Cues: Shape and Support
• Bee Syndrome: While not detailed extensively in these specific excerpts, the sources note that these flowers are designed for insects that often land to collect rewards like pollen or sucrose-dominated nectar.
• Moth Syndrome: Important moth pollinators, such as hawk moths, hover in front of flowers with rapid wingbeats. Consequently, moth flowers often have tubular corollas that accommodate the moth’s long tongue while it hovers, rather than providing a sturdy landing platform.
5. Reward Type and Volume
• Bee Syndrome: Plants offer sucrose-dominated nectar, pollen, or both as rewards. Some specialized flowers even provide oils from dedicated glands.
• Moth Syndrome: Because hovering flight is metabolically expensive, moth-pollinated flowers produce copious amounts of nectar to provide the high energy needed to fuel the moths’ high metabolic rates.
32. How do plants resolve the trade-off between attraction and defense?
Plants resolve the trade-off between attracting pollinators and defending against herbivores through a combination of mating system transitions, plastic defense syndromes, and structural adaptations that decouple the benefits of being “showy” from its genetic and ecological costs.
Plants utilize the following strategies to manage this conflict:
1. Mating System-Mediated Defense Strategies
The transition from outcrossing to self-fertilization fundamentally shifts a plant’s defensive syndrome.
• Constitutive vs. Inducible Defense: Outcrossing plants, which are often self-incompatible (SI), tend to rely on constitutive defenses (always active). This prevents the need for reactive chemical changes that might deter pollinators.
• The Advantage of Selfers: Inbreeding or self-compatible (SC) taxa exhibit resistance that is, on average, 63% more inducible (triggered only after attack) than their outcrossing relatives. Because selfing plants are less dependent on pollinators for every reproductive event—a benefit known as reproductive assurance—they have more evolutionary flexibility to deploy deterrent chemical defenses that would be too costly for outcrossers to maintain.
• Increased Specificity: Self-compatible species show higher specificity in their defenses, evolving independent responses to distinguish between generic mechanical wounding and actual herbivore damage. This allows them to “fine-tune” their defenses to minimize wasted resources while protecting reproductive tissues.
2. Structural Solutions to the “Cost of Being Showy”
Large floral displays are necessary to attract pollinators in competitive environments, but they lead to geitonogamy (pollen transfer between flowers on the same plant), which causes pollen discounting and inbreeding depression. Plants resolve this through:
• Spatial and Temporal Separation: Mechanisms like herkogamy (spatial separation of sex organs) and dichogamy (temporal separation) allow plants to maintain large, attractive displays while reducing the “mating costs” of geitonogamy.
• Dioecy (Separate Sexes): By evolving into separate male and female individuals, species can produce massive floral displays to maximize attraction with zero risk of self-pollination. This is especially common in long-lived or large plants where the risk of geitonogamy is highest.
3. Managing Chemical Conflicts in Reward Tissues
Pollinators and herbivores often interact with the same tissues, creating a conflict in floral chemistry.
• Pollinator Deterrence: The induction of toxic metabolites in nectar or pollen can repel the very pollinators a plant needs for outcrossing.
• Defense Plasticity: Plants use inducible defenses to resolve this, remaining “undefended” (and thus attractive) until an attack is confirmed. This saves the resource costs of defense and ensures that rewards remain palatable to mutualists until the threat of herbivory outweighs the benefit of attraction.
4. Specialized Co-evolutionary Compromises
In extreme cases, the plant resolves the trade-off by entering into antagonistic mutualisms where the pollinator and herbivore are the same species, such as yucca moths or fig wasps. In these systems, the plant “pays” for pollination by allowing the insect’s larvae to consume a portion of its seeds, essentially treating defense as a regulated cost of reproduction.
Summary of Trade-off Resolutions
| Conflict | Resolution Strategy | Functional Benefit |
|---|---|---|
| Pollinator vs. Herbivore Attraction | Inducible Defenses | Saves resources and maintains attraction until attack. |
| Floral Display vs. Selfing Cost | Herkogamy / Dichogamy | Minimizes geitonogamy while remaining showy. |
| Defense Toxin vs. Reward Quality | Defense Specificity | Prevents deterring pollinators while targeting herbivores. |
| Mate Scarcity vs. Seed Quality | Mixed Mating / Delayed Selfing | Prioritizes outcrossing but “fails safe” to selfing if needed. |
33. What is synchronized protandry in vertical flower spikes?
Synchronized protandry is a common form of dichogamy (the temporal separation of male and female functions) where the entire vertical inflorescence acts as the reproductive unit. In this system, individual protandrous flowers—which function as males before transitioning to females—mature sequentially from the bottom to the top of the flower spike.
This strategy utilizes the foraging behavior of pollinators to maximize outcrossing success through the following mechanism:
Mechanism of Upward Foraging
• Sequential Maturing: At any given time, the lower, older flowers on the spike are in their female phase (receptive stigmas), while the upper, younger flowers are in their male phase (dispersing pollen).
• Pollinator Path: Pollinators, particularly bees, typically land at the bottom of an inflorescence and forage upwards.
• Effective Transfer: By starting at the bottom, the pollinator deposits outcross pollen (brought from a previous plant) onto the receptive female-phase flowers. As it moves toward the top, it picks up fresh pollen from the male-phase flowers to carry to the next plant.
Evolutionary Advantages
Synchronized protandry resolves a fundamental conflict between a plant’s need to be “showy” and its need to produce high-quality offspring:
• Reduction of Geitonogamy: By segregating male and female functions among different flowers on the same spike, the plant minimizes geitonogamy (the transfer of pollen between flowers on the same individual), which is genetically equivalent to selfing and often leads to inbreeding depression.
• Minimizing Pollen Discounting: It reduces pollen discounting—the loss of outcross siring success caused by self-pollination. Pollen that would otherwise be “wasted” on the plant’s own stigmas remains available for export to other individuals.
• Maintaining Large Displays: This system allows plants to maintain larger floral displays to attract more pollinators without paying the typical “mating costs” of geitonogamy and sexual interference.
Evidence of Success
Experimental studies on species like Eichhornia paniculata and Chamerion angustifolium have demonstrated that inflorescences with synchronized protandry sire significantly more offspring than “adichogamous” inflorescences (where male and female functions occur simultaneously), despite having the same total amount of pollen and ovules. This confirms that the strategy specifically enhances a plant’s paternal fitness by promoting more efficient pollen dispersal.
34. How do pollinators influence the evolution of plant defense strategies?
Pollinators influence the evolution of plant defense strategies primarily by mediating the plant’s mating system, which dictates how resources are allocated between permanent (constitutive) and reactive (inducible) defenses.
Pollinators shape these strategies through the following mechanisms:
1. Driving the Shift from Constitutive to Inducible Defense
The most significant influence identified in the sources is the link between pollinator-mediated mating systems and defense deployment. Research in the nightshade family (Solanaceae) demonstrates that the transition from obligate outcrossing (self-incompatible, or SI) to inbreeding (self-compatible, or SC) drives the evolution of inducibility.
• Constitutive Strategy (Outcrossers): Plants that rely heavily on pollinators for outcrossing tend to favor higher levels of constitutive resistance—defenses that are always present.
• Inducible Strategy (Selfers): When pollinators are scarce and plants transition to self-compatibility, they evolve a more inducible strategy, becoming on average 63% more inducible than their outcrossing relatives. This allows the plant to save the metabolic cost of defense until an attack is confirmed.
2. Resolving Conflicting Selection in Reward Tissues
Pollinators exert selective pressure on the chemical traits of flowers, which can conflict with the plant’s need to deter herbivores.
• Pollinator Deterrence: Many chemical defenses are toxic or deterrent to pollinators if expressed in reward tissues like pollen and nectar.
• Evolutionary Flexibility: Obligately outcrossing plants pay a disproportionately high cost if they deter pollinators with chemical defenses. Self-compatible plants, being less dependent on pollinators for reproductive assurance, have more flexibility to deploy deterrent or toxic metabolites that might otherwise drive away mutualists.
3. Adaptation to Marginal Habitats
The sources suggest that self-compatible taxa often persist in marginal habitats or range edges where pollinators and mates are limiting. These environments are characterized by variable herbivory, which favors the evolution of phenotypic plasticity and inducible defenses over constant, high-level constitutive protection.
4. Increasing Defense Specificity
Pollinator-mediated selection appears to enhance the precision of a plant’s response. The loss of self-incompatibility is associated with the evolution of increased specificity in induced resistance.
• Precision: Self-compatible taxa are better at evolving independent responses to different types of damage, such as distinguishing between mechanical wounding and actual herbivore consumption.
• Outcrossing Correlation: In contrast, outcrossing (SI) taxa show lower specificity, with a strong correlation between their responses to wounding and real herbivory.
5. Specialized Antagonistic Mutualisms
In some co-evolved systems, the pollinator and the herbivore are the same individual.
• Obligate Antagonists: Examples include the yucca and yucca moth or figs and fig wasps. In these relationships, the moth is an obligate pollinator, but its larvae act as antagonistic herbivores that consume the plant’s seeds. This forces the plant to evolve defensive strategies that balance the survival of its seeds with the necessity of the insect’s pollination services.
6. Protection of Reproductive Structures
The method of pollination also dictates physical defense structures. For instance, beetle-pollinated flowers often have ovaries that are well-protected from the biting mouthparts of their pollinators. Similarly, different species using the same pollinators may flower asynchronously to avoid “stigma gunking” (heterospecific pollen transfer), which is a form of niche partitioning that protects reproductive quality.
35. How do plants use floral design to avoid self-fertilization?
Plants utilize diverse floral designs and functional strategies to prevent self-fertilization (selfing), which can lead to reduced offspring fitness through inbreeding depression. These strategies include spatial and temporal separation of sex organs, anatomical barriers between flowers, and physiological recognition systems.
1. Spatial Separation (Herkogamy)
One of the most widespread structural strategies is herkogamy, the physical separation of anthers (male organs) and stigmas (female organs) within a flower.
• Approach Herkogamy: Stigmas are positioned above anthers to ensure they contact a pollinator first upon entry, which effectively captures outcross pollen before the pollinator picks up the flower’s own pollen.
• Reciprocal Herkogamy (Heterostyly): This polymorphism involves populations with two (distyly) or three (tristyly) floral morphs that possess reciprocal arrangements of anthers and stigmas to facilitate cross-pollen transfer between morphs.
• Enantiostyly: Known as “mirror-image flowers,” the style is deflected either to the left or right while the pollinating anther is on the opposite side, which helps segregate pollen onto different parts of a pollinator’s body.
2. Temporal Separation (Dichogamy)
Dichogamy prevents selfing by separating the timing of anther dehiscence and stigma receptivity.
• Protogyny: The female phase occurs before the male phase, providing a receptive period free from the flower’s own pollen.
• Protandry: The male phase occurs first; while it may be less effective at preventing intrafloral selfing than protogyny, it reduces sexual interference.
• Synchronized Protandry: In vertical flower spikes, flowers mature from bottom to top; pollinators typically forage upwards, depositing outcross pollen on female-phase flowers at the bottom before picking up fresh pollen from male-phase flowers at the top.
3. Anatomical and Sexual Systems
Plants use the distribution of sex organs among flowers or individuals to ensure outcrossing.
• Dioecy: Populations consist of entirely separate male and female individuals, making self-fertilization morphologically impossible.
• Monoecy: Male and female functions are separated into different flowers on the same plant. While this prevents selfing within a single flower, it does not guarantee the prevention of geitonogamy (selfing between different flowers on the same individual).
4. Physiological Rejection (Self-Incompatibility)
Many species possess self-incompatibility (SI), a genetic recognition system that allows a plant to reject its own pollen.
• Pre-zygotic Rejection: When a plant’s own pollen lands on its stigma, physiological processes inhibit pollen germination or pollen-tube growth, preventing fertilization entirely.
• Ovarian or Late-acting SI: Rejection occurs after the pollen tubes have already entered the ovary, often involving signaling that prevents embryo development.
5. Specialized Mechanical Responses
Some floral designs include rapid developmental or mechanical changes to avoid interference.
• Stigma Closure: Some species have sensitive stigmas that close upon mechanical stimulation from a pollinator; this can promote more effective outcross pollen dispersal and limit the risk of subsequent “stigma gunking” by self-pollen.
• Pollinia: Orchids package pollen into clusters called pollinia, which are “glued” to specific parts of an insect to ensure targeted transfer between individuals rather than accidental self-pollination.
36. Why do some plants exhibit different sexual systems across populations?
Plants exhibit different sexual systems across populations because of a complex interplay between environmental conditions, habitat stability, and metapopulation dynamics that shift the balance between the need for outcrossing and the necessity of reproductive assurance. While approximately 6% of angiosperm species are dioecious (separate sexes) and most others are hermaphroditic, some “labile” species maintain multiple sexual systems across their range.
The primary reasons for this variation include:
1. Habitat Stability and Disturbance
Variation in sexual systems often reflects differences in the longevity and stability of a population’s environment.
• Case Study (Sagittaria latifolia): This species possesses both monoecious (separate flowers on one plant) and dioecious populations. Monoecious populations primarily colonize ephemeral, frequently disturbed habitats. In these environments, populations are short-lived, and the capacity for geitonogamous selfing provides a vital “safety net” for reproduction.
• Dioecious Populations: Conversely, dioecious populations of S. latifolia inhabit large, stable marshlands where individuals can form massive, long-lived clones. Because large clones are highly susceptible to geitonogamy (selfing between flowers on the same plant), which causes severe inbreeding depression, separate sexes are selected for to enforce outcrossing.
2. Environmental Stress and Aridity
The physical environment, such as water availability, can dictate which sexual system is most viable.
• Case Study (Ecballium elaterium): In the Iberian Peninsula, monoecious populations of this perennial occur in wetter northern regions, but they are replaced by dioecious populations in the arid south.
• Stress-Induced Dimorphism: Arid or high-stress conditions may promote the evolution of separate sexes because cosexual plants may be unable to maintain both male and female functions when resources are limited. Alternatively, stress may magnify the costs of inbreeding depression, favoring the spread of unisexual individuals.
3. Metapopulation Dynamics and Baker’s Law
For species whose populations experience frequent cycles of colonization and extinction, the ability to self-fertilize is a major selective advantage.
• Reproductive Assurance: Consistent with Baker’s Law, individuals that can produce seeds without a mate are better colonizers. In the weed Mercurialis annua, populations vary between dioecious, monoecious, and androdioecious (males and cosexuals) across Europe.
• Selection Against Unisexuality: In short-lived, isolated colonies where mates are scarce, metapopulation processes select against unisexuality and self-incompatibility, favoring hermaphroditism or monoecy for reproductive assurance.
4. Size-Dependent Gender Modification
In some species, the sexual system varies across populations because gender expression is tied to plant size and resource availability.
• Case Study (Wurmbea dioica): This species exhibits a range of systems from cosexuality to full dioecy. In subdioecious populations, male plants are often labile, producing hermaphrodite flowers only when they grow large enough to bear the high energetic costs of seed and fruit development.
• Population Segregation: These cosexual and sexually dimorphic populations can exist within meters of each other, maintained as distinct units by differences in flowering time.
5. “Leaky” Sex Expression as a Fail-Safe
Even in strictly dioecious populations, plants may exhibit “leaky” sex expression—where a unisexual plant produces flowers of the opposite sex—in response to local mate scarcity.
• Experimental Evidence: In an experiment with Mercurialis annua, the systematic removal of males from a dioecious population led to females evolving male function to reproduce via self-fertilization. This suggests that what appears to be a stable sexual system in one population can rapidly transition if the demographic environment changes.
37. How do plants adjust their gender based on their size?
Plants adjust their gender based on their size through a process called size-dependent gender modification, where an individual plant shifts its relative investment in male and female functions as it grows and accumulates resources. This flexibility is possible because plants have a modular construction that allows them to reiterate reproductive units in different spatial and temporal combinations.
The primary ways plants manage this adjustment include:
1. The Energetic Cost of “Motherhood”
The most common pattern is for plants to increase their relative allocation to female function as they grow larger.
• Energetic Demands: Producing seeds and fruits is significantly more energetically expensive than producing pollen.
• Growth Thresholds: Because of these high costs, many plants begin reproducing as males when they are small or young. They only develop female functions—either by becoming hermaphroditic or transitioning entirely to a female phase—once they have reached a size threshold capable of supporting maternal investment.
2. Case Study: Wurmbea dioica
In subdioecious populations of the Australian geophyte Wurmbea dioica, gender expression is highly labile in response to size.
• Inconstant Males: Male plants in these populations often produce only male flowers when small.
• Hermaphroditic Shift: As these plants grow larger and their resource-storing corms expand, they begin to produce hermaphrodite flowers, adding female function to their existing male function because they can now bear the reproductive costs of developing seeds.
3. The Wind-Pollination Exception
While animal-pollinated plants generally become more “female” as they grow, some wind-pollinated species show the opposite trend, becoming more male-biased as they get taller.
• Dispersal Efficiency: For wind-pollinated plants (such as those in the Poaceae or Asteraceae families), being taller provides a distinct advantage for male fitness.
• Height Advantage: Taller individuals can disperse their pollen over much wider areas via wind currents, leading to higher fitness returns from male investment compared to smaller individuals of the same species.
4. Environmental Interactions
The sources note that size-dependent gender adjustment is often a response to environmental quality. In “poor” or low-resource environments, plants are more likely to remain or function as males, as female fertility appears more susceptible to limitation by low environmental quality than male fertility. Female function is typically reserved for “good” environments where the plant has the biomass and resources to ensure the survival of its seeds.
38. What are the common rewards plants offer to attract pollinators?
Plants offer a variety of metabolic and physical rewards to pollinators to ensure repeated visits, which are critical for the transfer of pollen between individuals. While floral traits like color and scent function as “attraction strategies” to signal the presence of these resources, the rewards themselves are the primary incentive for the pollinator’s behavior.
The most common rewards include:
1. Nectar
Nectar is the most widespread reward offered by flowering plants. Its volume and chemical composition are often tailored to the specific metabolic needs of the target pollinator:
• Bees: Often receive sucrose-dominated nectar in varying amounts.
• Moths and Birds: These high-energy foragers require copious amounts of nectar to fuel the metabolic costs of hovering flight.
• Butterflies: Since they generally do not digest pollen, they rely almost exclusively on nectar, which they reach using their long proboscis.
• Specialist Nectarivores (Birds): Typically receive large volumes of dilute nectar secreted during the day.
2. Pollen
Pollen serves a dual purpose as both the plant’s male gametes and a protein-rich food source for many insects.
• Edible Pollen: Many plants produce pollen specifically meant for consumption by pollinators like bees and beetles.
• Feeding Anthers: In species with enantiostyly (mirror-image flowers), there is often a functional division of labor where “feeding anthers” provide pollen as a reward, while separate “pollinating anthers” handle the actual transfer of genetic material.
• Pollinia: Some specialized plants like orchids package pollen into clusters called pollinia; while not always a food reward, they are “glued” to the insect during its attempt to access other rewards or through mimicry.
3. Oils
Specialized floral glands produce oils as a reward for a specific group of “oil-collecting” bees. This is a more specialized reward than nectar or pollen and is found in a relatively small number of plant species.
4. Heat
In some cases, heat is considered a reward or a specialized means of attraction.
• Thermogenic Flowers: Certain plants, such as those pollinated by beetles, can increase their internal temperature.
• Functional Benefits: This heat can help further spread floral scents, act as an infrared “shining beacon” to attract insects in the dark, or provide a warm refuge for pollinators in cold environments.
5. Deceptive Strategies (No Reward)
It is important to note that not all plants provide a reward. Sapromyophilous plants attract flies by mimicking the odor of dead animals or dung but provide no reward. Similarly, some specialized orchids use mimicry to resemble female insects, tricking males into attempting to mate with the flower without receiving any metabolic benefit.
39. Are there plants that mimic insects to trick pollinators?
Yes, there are plants that utilize highly specialized mimicry to trick pollinators into visiting them, a strategy most notably found within the orchid family (Orchidaceae),. These plants employ both visual and chemical deceptions to attract specific insects for pollination.
These mimicry strategies include:
• Morphological Mimicry: Some orchids have evolved flower shapes that precisely mimic the physical appearance of insects. This structural resemblance is designed to trick male insects into attempting to “mate” with the flower.
• Chemical Mimicry: In addition to visual cues, a few specialized orchids produce scents that mimic insect pheromones. These chemical signals are often indistinguishable from those produced by sexually receptive female insects, drawing in males from a distance.
• Specific Example (The Bee Orchid): The bee orchid (Ophrys apifera) is a classic example of this phenomenon. It has evolved to look and smell like a sexually receptive female bee, which attracts naïve male bees who inadvertently pollinate the flower while attempting to mate with it.
These “deceptive” plants typically provide no metabolic reward, such as nectar or edible pollen, to the pollinator. Instead, they rely entirely on the biological drive of the insect to find a mate to ensure the transfer of their pollinia (pollen clusters) to the next flower.
40. Why do some plants choose to offer no reward at all?
Some plant species, most notably within the orchid family (Orchidaceae), utilize deceptive pollination strategies that offer no metabolic reward (such as nectar or edible pollen) to their pollinators. This approach allows the plant to achieve reproduction while avoiding the energetic costs of producing resources for animal visitors.
Plants choose this “no-reward” strategy for the following reasons and through the following mechanisms:
1. Exploiting Biological Drives (Sexual Mimicry)
Plants can achieve pollination without offering food by exploiting an insect’s drive to find a mate.
• Morphological and Chemical Mimicry: Many specialized orchids have evolved flower shapes that mimic the physical appearance of insects. Some even produce scents that mimic insect pheromones.
• Pseudocopulation: A classic example is the bee orchid (Ophrys apifera), which resembles and smells like a sexually receptive female bee. It attracts naïve male bees that inadvertently pollinate the flower while attempting to mate with it, receiving no reward for their efforts.
2. Mimicking Brood Sites or Food (Sapromyophily)
Some plants attract pollinators by mimicking undesirable substances that certain insects normally visit to lay eggs or feed.
• Sapromyophilous Plants: These plants attract flies by mimicking the odor of dead animals or dung.
• Brief Interactions: Because the flower provides no actual food, the flies typically leave quickly. However, the plant may employ traps to slow the insects down, ensuring enough time for pollen transfer before the insect realizes it has been deceived.
3. Reducing Energetic and Genetic Costs
Choosing not to offer a reward can be a way for a plant to manage its resource allocation efficiently.
• Attraction without Investment: Flowers are essentially “attraction strategies”. By using visual or olfactory “false advertising,” a plant can maximize its visual effect and noticeability to passing pollinators while saving the resources that would otherwise go into nectar glands or edible pollen.
• Sterile Flowers: In some cases, the evolution of sterile flowers can reduce pollen limitation by increasing the overall attraction of an inflorescence at a very low energetic and genetic cost to the plant.
4. Pollination for Non-Genetic Reasons (Pseudogamy)
In some asexual reproductive systems, a plant may require pollination even if the pollen provides no genetic benefit to the offspring.
• Triggering Embryo Growth: In a process called pseudogamy, pollination is often necessary to initiate embryo growth in apomictic (asexual) seeds.
• No Genetic Reward: Even though the pollinator carries pollen to the plant, that pollen contributes no genetic material to the developing offspring, representing a biological interaction where the “reward” of genetic recombination is absent.
In summary, plants offer no reward as a specialized evolutionary compromise. By mimicking mates, food, or brood sites, they exploit the sensory systems and behaviors of specific pollinators to ensure gene transfer while minimizing the metabolic expense of reward production.
41. How do plants ensure cross-pollination in large clonal displays?
Plants ensure cross-pollination in large clonal displays by evolving specialized floral designs and growth architectures that decouple the benefits of being showy (attracting pollinators) from the genetic costs of geitonogamy (pollen transfer between flowers on the same individual or clone). Because geitonogamy is genetically equivalent to self-fertilization, it often leads to severe inbreeding depression and “pollen discounting”—the loss of outcross siring success.
Several key mechanisms plants use to promote cross-pollination in these contexts:
1. Structural and Temporal Separation
Plants utilize structural modifications within their flowers and inflorescences to manage how pollinators interact with male and female sex organs.
• Herkogamy (Spatial Separation): By separating anthers and stigmas in space, plants contact pollinators in a specific sequence that captures outcross pollen before picking up the plant’s own pollen.
• Dichogamy (Temporal Separation): By separating the timing of male and female function, plants reduce the likelihood that a pollinator will transfer pollen between flowers on the same individual.
• Synchronized Protandry: In vertical flower spikes, flowers mature from the bottom up. Lower, older flowers are in the female phase, while upper, younger flowers are in the male phase. Because pollinators like bees typically forage upwards, they deposit outcross pollen from other plants on the bottom flowers and pick up fresh pollen from the top flowers to carry away, effectively bypassing the risk of self-pollination within the spike.
2. Transitions to Separate Sexes (Dioecy)
In long-lived species that form massive clones, the risk of geitonogamy is so high that it can destabilize hermaphroditism.
• Selective Enforcement of Outcrossing: Evolution often favors dioecy (separate male and female individuals) in clonal populations. In these species, individuals can produce enormous floral displays for attraction with zero risk of self-fertilization, as each individual only possesses one sex.
• Case Study (Sagittaria latifolia): This aquatic plant maintains monoecious populations (separate flowers on one plant) in unstable habitats where clones stay small, but transitions to dioecy in stable marshlands where clones become large enough that geitonogamous selfing would otherwise exceed 50%.
3. Clonal Architecture and Spatial Mixing
The physical arrangement of a clone’s growth influences mating opportunities:
• Guerilla vs. Phalanx Strategies: Some clonal plants use “guerilla” strategies or have fragmentary growth with weak stolon connections. This allows the daughter rosettes of a single genotype to spread and intermingle with different genotypes, reducing the probability that a pollinator will visit only a single clone during its foraging bout.
• Temporal Asynchrony: In extensive clonal colonies where shoots are spatially segregated but physiologically autonomous, a high degree of temporal asynchrony between male and female sex function across the entire clone is required to prevent selfing.
4. Physiological Barriers
When structural strategies fail to prevent self-pollen deposition, plants rely on physiological systems:
• Self-Incompatibility (SI): Many showy plants possess SI systems that recognize and reject self-pollen grains if they land on the stigma. This ensures that even if a pollinator visits many flowers on the same clone, only pollen from a different individual can successfully fertilize the ovules.
• Cryptic Self-Incompatibility: Some species use “pollen-pistil interactions” where outcross pollen is given prepoterance (higher speed) over self-pollen, ensuring that if both land on a stigma, the outcross pollen wins the “race” to the ovule.
5. Management of Pollen Resources
Large clonal displays are prone to pollen discounting, where self-pollination “wastes” pollen that could have been used to sire seeds on other plants. Strategies like heterostyly (reciprocal arrangements of sex organs) function to increase the proficiency of pollen dispersal by placing pollen on specific parts of a pollinator’s body that will only contact the stigmas of a different floral morph, thereby reducing interference and promoting outcrossing.
42. How do plants separate the showy display from selfing costs?
Plants separate the benefits of a showy floral display (attracting pollinators) from the genetic and metabolic costs of self-fertilization (selfing) through a variety of structural, temporal, and physiological mechanisms known as mating strategies.
While large floral displays are necessary to attract pollinators, they increase the likelihood of geitonogamy—the transfer of pollen between flowers on the same individual—which is genetically equivalent to selfing and carries high costs, including inbreeding depression and pollen discounting (the loss of outcross siring success).
Plants resolve these conflicts using the following strategies:
1. Structural Separation (Herkogamy)
Plants use herkogamy, the physical separation of anthers and stigmas, to manage how pollinators interact with their sex organs.
• Approach Herkogamy: Stigmas are positioned above anthers so they contact the pollinator first upon entry, capturing outcross pollen before the pollinator picks up the flower’s own pollen.
• Reciprocal Herkogamy (Heterostyly/Enantiostyly): These polymorphisms involve Complimentary floral morphs with reciprocal arrangements of sex organs (e.g., “mirror-image” flowers). These designs increase the proficiency of cross-pollen transfer and reduce male gamete wastage caused by sexual interference.
2. Temporal Separation (Dichogamy)
Dichogamy separates the timing of male and female functions within or between flowers.
• Protogyny: The female phase occurs before the male phase, providing a receptive period free from the flower’s own pollen.
• Synchronized Protandry: In vertical flower spikes, flowers mature from the bottom to the top. Because pollinators like bees typically forage upwards, they deposit outcross pollen on receptive female flowers at the bottom and pick up fresh pollen from male flowers at the top to carry to another plant. This allows plants to maintain larger floral displays without paying the mating costs associated with geitonogamy.
3. Transitions to Separate Sexes (Dioecy)
In long-lived plants or those that form massive clones, the risk of geitonogamy is so high that hermaphroditism can become maladaptive.
• Enforced Outcrossing: Selection often drives the evolution of dioecy (separate male and female individuals). Dioecious plants can produce enormous, showy displays to maximize attraction with zero risk of self-fertilization.
• Habitat Stability: Research on Sagittaria latifolia shows that populations in stable marshlands transition to dioecy to bypass the massive geitonogamy costs that occur when clones grow large.
4. Physiological Rejection (Self-Incompatibility)
Many showy plants possess self-incompatibility (SI), a genetic recognition system that rejects a plant’s own pollen.
• Gametophytic/Sporophytic SI: These systems inhibit pollen germination or tube growth if the pollen matches the maternal genotype.
• Late-Acting SI: Some species, like those in the genus Narcissus, use ovarian or late-acting SI, rejecting self-pollen after it has already reached the ovary.
• Cryptic SI: This involves “pollen-pistil interactions” where outcross pollen is given prepotency (higher growth speed) over self-pollen, ensuring outcrossing even if a mixture of pollen is present.
5. Clonal Growth Architectures
Some plants manage geitonogamy costs through their growth form. Fragmentary growth (guerilla strategies) allows daughter rosettes of a single genotype to spread and intermingle with different genotypes, reducing the probability that a pollinator will visit only a single clone during a foraging bout.
43. How do pollinators influence the evolution of plant defense strategies?
Pollinators influence the evolution of plant defense strategies primarily by mediating the plant’s mating system, which determines the costs and benefits of deploying permanent (constitutive) versus reactive (inducible) defenses.
The relationship between pollinators and defense evolution is shaped by the following factors:
1. Driving the Shift from Constitutive to Inducible Defense
The transition from obligate outcrossing (facilitated by pollinators) to self-compatibility (which allows for inbreeding) drives a major macroevolutionary shift in defense deployment.
• Constitutive Strategy in Outcrossers: Plants that rely heavily on pollinators for outcrossing (self-incompatible species) tend to favor constitutive defenses—those always present regardless of attack.
• Inducible Strategy in Selfers: In contrast, plant resistance is on average 63% more inducible in self-compatible taxa than in their outcrossing relatives. This shift allows plants to save the metabolic cost of defense until an actual attack occurs.
2. The “Pollinator Deterrence” Constraint
Pollinators act as a selective constraint on chemical defenses, particularly in “reward tissues” like nectar and pollen.
• Cost of Deterrence: For obligately outcrossing plants, inducing toxic or deterrent metabolites can be disproportionately costly because it may drive away the very pollinators they require for reproduction.
• Evolutionary Flexibility: Because self-compatible plants are less dependent on pollinators for every reproductive event—a benefit known as reproductive assurance—they have more evolutionary flexibility to deploy deterrent chemical defenses that would be too risky for outcrossers.
3. Increasing Defense Specificity
The loss of reliance on pollinators (transition to selfing) is associated with the evolution of increased specificity in defenses.
• Precision: Self-compatible taxa are better at “fine-tuning” their responses to distinguish between generic mechanical wounding and actual herbivore damage.
• Lack of Independence in Outcrossers: Outcrossing (self-incompatible) taxa show lower specificity, with their responses to wounding and real herbivory evolving dependently.
4. Adaptation to Marginal Habitats
Self-compatible species often persist in marginal habitats or range edges where pollinators and mates are scarce.
• Variable Herbivory: These environments are often characterized by variable herbivory, which theoretical models predict will favor the evolution of phenotypic plasticity and inducible strategies over constant high-level defense.
5. Specialized Antagonistic Mutualisms
In some highly co-evolved systems, the pollinator and the herbivore are the same species, creating a direct conflict in defense evolution.
• Obligate Antagonists: In relationships like the yucca and yucca moth or figs and fig wasps, the insect is an obligate pollinator, but its larvae are antagonistic herbivores that consume the plant’s seeds.
• Regulated Costs: In these cases, the plant’s defensive strategy must balance the survival of its seeds with the necessity of the insect’s pollination services, essentially treating seed consumption as a regulated cost of reproduction.
6. Physical Protections
Pollinator foraging behavior also dictates the evolution of physical defense structures. For example:
• Beetle Pollination: Because beetles have biting mouthparts that can damage reproductive tissues, beetle-pollinated flowers often evolve ovaries that are well protected or buried deep within the flower.
• Niche Partitioning: Different plant species sharing the same pollinators may evolve to flower asynchronously to avoid “stigma gunking” (heterospecific pollen transfer), a form of reproductive defense that protects the quality of the mating event.
44. How do plants like orchids use sexual mimicry to deceive?
Plants in the orchid family (Orchidaceae) frequently use highly specialized sexual mimicry as a deceptive pollination strategy to attract specific insects without providing any metabolic reward. This deception targets male insects by exploiting their biological drive to find a mate.
Orchids achieve this through two primary modes of deception:
• Morphological (Visual) Mimicry: Many orchids have evolved floral shapes and structures that precisely mimic the physical appearance of an insect to trick males into thinking they have found a partner.
• Chemical Mimicry: In addition to visual cues, some orchids produce scents that mimic insect pheromones. These chemical signals draw male insects from a distance, even when the flower is not yet visible.
The Mechanism of Pseudocopulation
A classic example provided in the sources is the bee orchid (Ophrys apifera), which has evolved to visually resemble and olfactorily smell like a sexually receptive female bee. When a naïve male bee is lured into attempting to “mate” with the flower—a process often referred to as pseudocopulation—it is forced into a specific position by the flower’s shape.
During this interaction, specialized clusters of pollen called pollinia become “glued” or attached to the insect’s body as it attempts to engage with the flower. As the male insect travels to another orchid to repeat the behavior, it inadvertently facilitates targeted cross-pollination.
Evolutionary Advantages
By using these deceptive attraction strategies, orchids are able to ensure consistent pollination services while bypassing the energetic and metabolic costs of producing rewards such as nectar or edible pollen. This allows the plant to function as a highly efficient “attraction strategy,” preserving its resources for other functions like seed production.
45. What are the survival benefits of offering no reward to pollinators?
Offering no reward to pollinators is a specialized evolutionary compromise that allows plants to achieve reproductive success while maximizing metabolic efficiency and resource allocation.
The primary survival benefits of this “no-reward” or deceptive strategy include:
1. Resource Conservation and Metabolic Efficiency
Producing rewards like nectar or large quantities of edible pollen is energetically expensive. For example, moth-pollinated flowers must produce copious amounts of nectar to fuel the high metabolic rates required for the insects’ hovering flight. By utilizing deceptive attraction—such as mimicking the scent of carrion or the appearance of a mate—a plant can bypass these production costs entirely, preserving its energy for other vital functions like seed development or growth.
2. Reduced Pollen Limitation at Low Cost
Plants can increase the overall visibility and attraction of their displays without a proportional increase in resource expenditure.
• Sterile Flowers: Some species evolve sterile flowers within an inflorescence. These act as additional visual lures to reduce pollen limitation by increasing the attraction of pollinators at a very low energetic and genetic cost to the plant.
• Visual and Chemical “False Advertising”: Many orchids use highly specialized flower shapes or scents that mimic insect pheromones to attract specific pollinators. This allows the plant to act as an efficient “attraction strategy” that ensures consistent pollination services without having to “pay” the pollinator with food.
3. Strategic Manipulation of Pollinator Behavior
Deceptive plants often employ mechanisms to ensure that even a brief, unrewarded visit results in successful pollen transfer.
• Traps: Because pollinators realize they have been deceived and leave “no-reward” flowers quickly, some plants, such as sapromyophilous species (which mimic dung or dead animals) or beetle-pollinated species, use traps. These physical structures slow the pollinator down or temporarily confine it, ensuring sufficient time for pollen to be attached or deposited before the insect escapes.
• Precise Pollen Packaging: Orchids package their pollen into clusters called pollinia, which are “glued” to the insect during its attempt to mate with or investigate the flower. This ensures that even a single deceptive encounter can lead to the transfer of thousands of pollen grains to the next plant.
4. Niche Partitioning and Specificity
By mimicking very specific triggers, such as the unique pheromones of a single insect species, plants can achieve a high degree of pollination specialization. This specialization can be a survival advantage because it leads to more consistent and targeted pollination services, reducing the risk of “stigma gunking” (the transfer of pollen from the wrong species).
References
Ballesteros-Mejia, L., Lima, N. E., Lima-Ribeiro, M. S., & Collevatti, R. G. (2016). Pollination mode and mating system explain patterns in genetic differentiation in neotropical plants. PLoS ONE, 11(7). https://doi.org/10.1371/journal.pone.0158660
Barrett, S. C. H. (1998). The evolution of mating strategies in flowering plants (Vol. 3, Number 9).
Barrett, S. C. H., Richards, A. J., Bayliss, M. W., Charlesworth, D., & Abbott, R. J. (2003). Mating strategies in flowering plants: The outcrossing-selfing paradigm and beyond. Philosophical Transactions of the Royal Society B: Biological Sciences, 358(1434), 991–1004. https://doi.org/10.1098/rstb.2003.1301
Campbell, S. A., & Kessler, A. (2013). Plant mating system transitions drive the macroevolution of defense strategies. Proceedings of the National Academy of Sciences of the United States of America, 110(10), 3973–3978. https://doi.org/10.1073/pnas.1213867110
Charlesworth, D. (2006). Evolution of Plant Breeding Systems. In Current Biology (Vol. 16, Number 17). https://doi.org/10.1016/j.cub.2006.07.068
Clo, J., Awad, D. A., Bilde, T., Bocedi, G., Haag, C. R., Pannell, J., & Hartfield, M. (2025). Perspectives on mating–system evolution: comparing concepts in plants and animals. In Journal of Evolutionary Biology (Vol. 38, Number 6, pp. 673–692). Oxford University Press. https://doi.org/10.1093/jeb/voaf009
Devaux, C., Porcher, E., & Lande, R. (2019). Mating systems and avoidance of inbreeding depression as evolutionary drivers of pollen limitation in animal-pollinated self-compatible plants. Annals of Botany, 123(2), 327–336. https://doi.org/10.1093/aob/mcy181
Eckert, C. G., Kalisz, S., Geber, M. A., Sargent, R., Elle, E., Cheptou, P. O., Goodwillie, C., Johnston, M. O., Kelly, J. K., Moeller, D. A., Porcher, E., Ree, R. H., Vallejo-Marín, M., & Winn, A. A. (2010). Plant mating systems in a changing world. In Trends in Ecology and Evolution (Vol. 25, Number 1, pp. 35–43). https://doi.org/10.1016/j.tree.2009.06.013
Meyer, E. M., Galloway, L. F., & Eckert, A. J. (2025). The evolutionary dynamics of plant mating systems: how bias for studying ‘interesting’ plant reproductive systems could backfire. Annals of Botany, 136(1), 15–27. https://doi.org/10.1093/aob/mcaf031
Suijkerbuijk, H. A. C., Ramos, S. E., & Poelman, E. H. (2025). Plasticity in plant mating systems. In Trends in Plant Science (Vol. 30, Number 4, pp. 424–436). Elsevier Ltd. https://doi.org/10.1016/j.tplants.2024.10.013
Whitehead, M. R., Lanfear, R., Mitchell, R. J., & Karron, J. D. (2018). Plant mating systems often vary widely among populations. In Frontiers in Ecology and Evolution (Vol. 6, Number APR). Frontiers Media S. A. https://doi.org/10.3389/fevo.2018.00038
Related posts:
 The Evolutionary Genomics of Crop Plants: From Ancestral Polyploidy to Modern Pangenomes
The Evolutionary Genomics of Crop Plants: From Ancestral Polyploidy to Modern Pangenomes
 Global Strategies for Crop Germplasm Conservation: Integrating Farmer-Based In Situ Methods with Biotechnological Innovation
Global Strategies for Crop Germplasm Conservation: Integrating Farmer-Based In Situ Methods with Biotechnological Innovation
 The Architecture of Agricultural Resilience: A Definitive Guide to Centers of Origin and Genetic Diversity
The Architecture of Agricultural Resilience: A Definitive Guide to Centers of Origin and Genetic Diversity
 ढलौटे फोहोरको बायोचार र Trichoderma afroharzianum सँग मिलेर टमाटरमा हुने जराको रोग नियन्त्रण
ढलौटे फोहोरको बायोचार र Trichoderma afroharzianum सँग मिलेर टमाटरमा हुने जराको रोग नियन्त्रण
 Modern Plant Breeding: Accelerating Genetic Gain and Climate Resilience through Integrated Genomic Selection and Synthetic Biotechnology
Modern Plant Breeding: Accelerating Genetic Gain and Climate Resilience through Integrated Genomic Selection and Synthetic Biotechnology