1. Introduction: The Strategic Imperative of Genetic Diversification
Modern agriculture is, in many ways, a victim of its own success. The “Green Revolution” focused on high-yielding, uniform cultivars, but this pursuit of consistency created a genetic bottleneck. By narrowing the genetic base, we inadvertently discarded the rare and favorable allelic variations that allowed wheat’s ancestors to survive for millennia without human intervention.
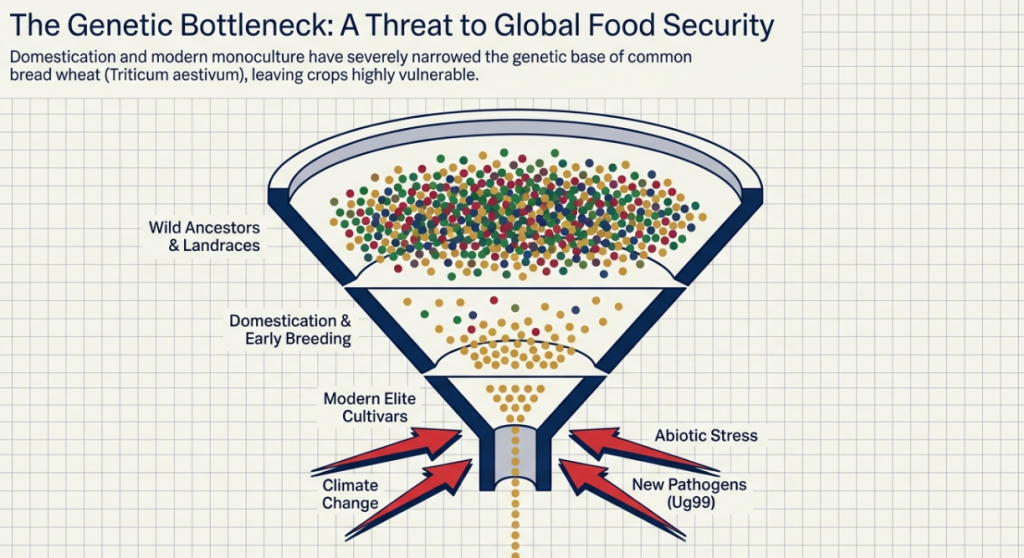
This loss of diversity has led to a “Hidden Hunger” crisis—a global deficiency in vital micronutrients like Iron (Fe) and Zinc (Zn) that affects over three billion people. To move forward, one must learn to see the limits of our current crops through the lens of ancestral potential.
The Pillar of Human Nutrition: Why Wheat Matters
Wheat stands as the foundational titan of global food security. As a Senior Agricultural Bio-Scientist, I view wheat not merely as a crop, but as the primary biological vehicle for delivering energy and essential nitrogenous compounds to the human species. In global agriculture, two species represent nearly the entirety of production: Bread Wheat (Triticum aestivum L.), a hexaploid species comprising approximately 95% of cultivation, and Durum Wheat (Triticum durum L.), a tetraploid species accounting for the remaining 5% and serving as the gold standard for pasta and semolina.
The nutritional mandate of wheat is evidenced by its staggering contribution to the global diet:
- 20% of Global Calories: Wheat provides one-fifth of the total energy required by the human population.
- 20% of Global Proteins: Remarkably, wheat contributes more to total human protein intake than the global consumption of all meat sources combined.
Despite its current dominance, the narrow genetic base of modern cultivars leaves our food supply vulnerable to climate volatility and emerging pathogens. To secure the future, the breeder’s mandate is to exploit the untapped genetic reservoirs found in wheat’s ancestral past.
Modern Wheat vs. Ancestral Potential
| Feature | Modern Wheat Cultivars | Ancestral & Wild Relatives |
|---|---|---|
| Genetic Diversity | Narrow (Uniformity/Bottleneck) | Wide (Rare Allelic Variation) |
| Yield Strategy | Optimized for high-input/stable environments | Variable, but high natural resilience |
| Stress Resistance | Vulnerable to shifting biotic/abiotic stress | “Superpowers” against pests and diseases |
| Nutritional Profile | Often lower in Fe and Zn (Hidden Hunger) | Unique sources of high mineral density |
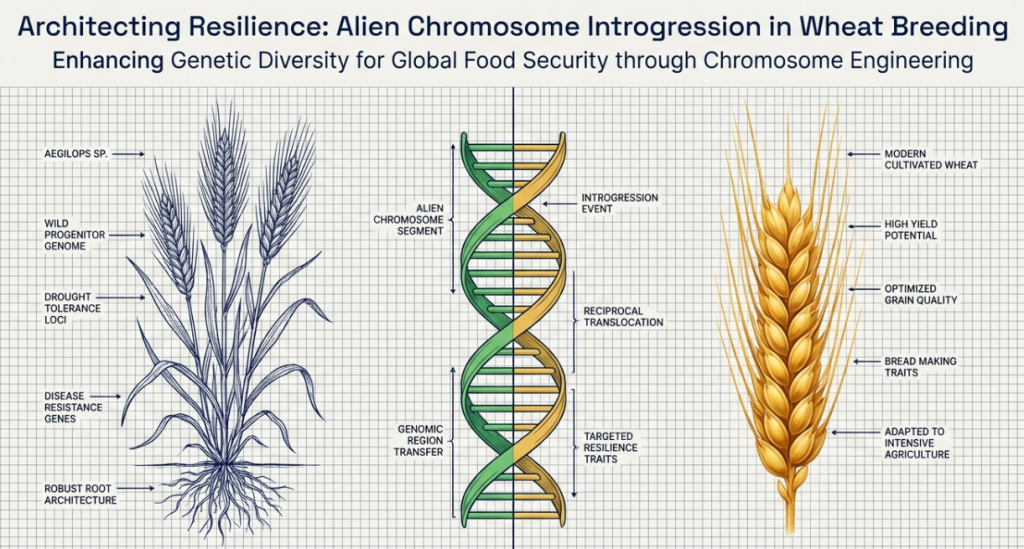
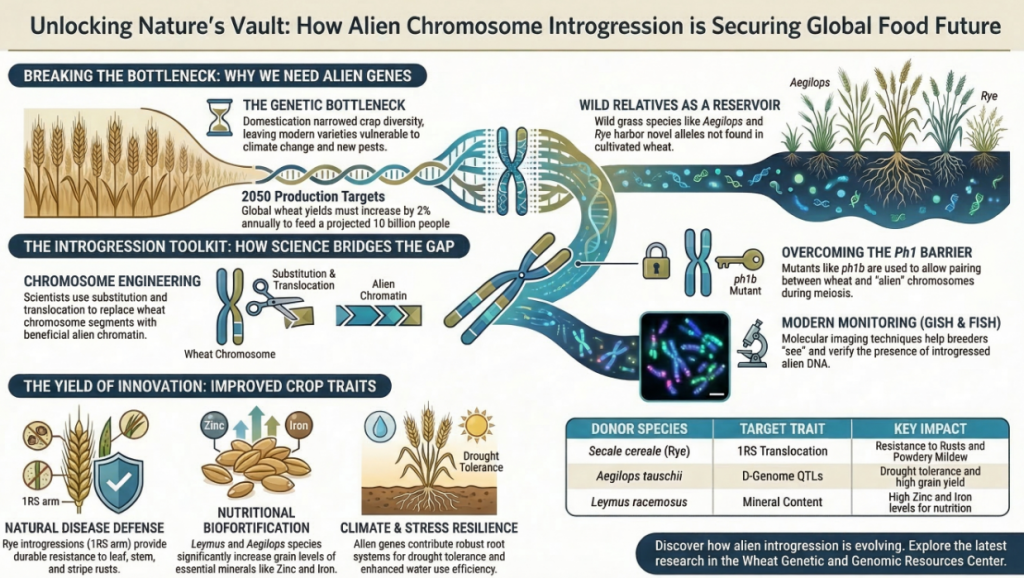
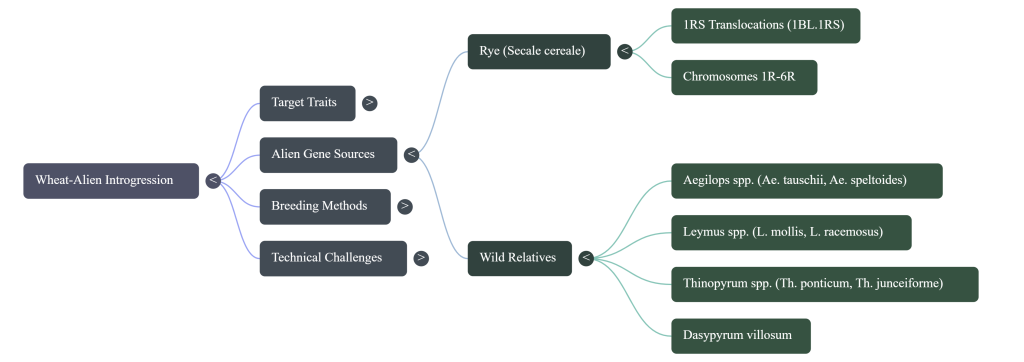
To achieve sustainable agriculture, we must look beyond the exhausted primary gene pool and strategically expand into the secondary and tertiary gene pools of the Triticeae tribe. Alien Chromosome Introgression is the primary cytogenetic methodology to restore this favorable allelic variation. By utilizing wild relatives to “mine” and incorporate novel genetic segments, we can circumvent current yield plateaus and address the “hidden hunger” of micronutrient deficiencies, ensuring global food security through 2050. The strategic value of these introgressions lies in their capacity to restore alleles lost during the polyploidization bottleneck, providing a mechanism to overcome current limitations regarding yield stability and nutrient density. Wild relatives—specifically Secale cereale (rye), Leymus spp., and the Thinopyrum genus—serve as indispensable reservoirs of novel alleles.
Defining “Alien Introgression”: Biology’s Version of a System Upgrade
Alien Introgression is the elegant process of transferring favorable alleles from “alien” species—those outside the immediate Triticum genus—into our cultivated bread wheat. Imagine this as a biological system upgrade: we are borrowing “source code” from distant cousins to patch the vulnerabilities in our current genetic operating system.
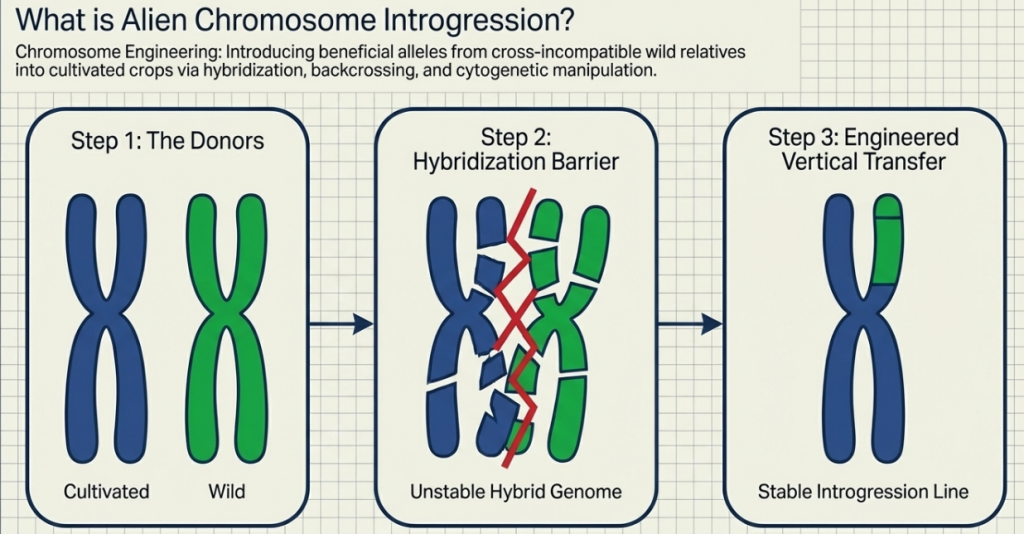
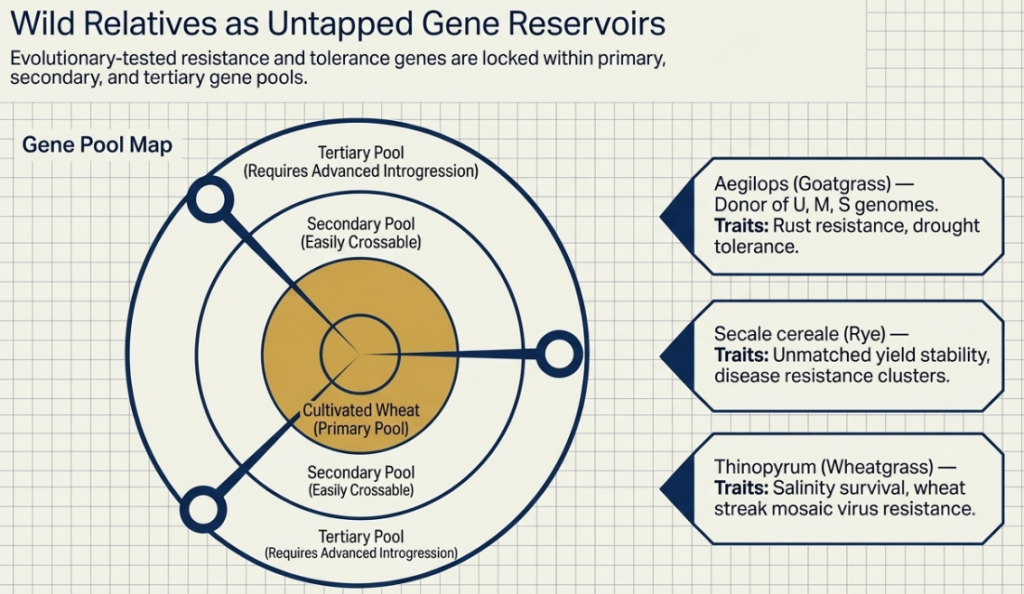
The difficulty of this upgrade depends on the “Gene Pool” of the wild relative:
- Primary Gene Pool: Direct ancestors (like Ae. squarrosa). These cross easily with wheat because they share homologous genomes.
- Secondary Gene Pool: Species sharing at least one genome with wheat (like Ae. cylindrica). Transfers are possible but require sophisticated selection.
- Tertiary Gene Pool: Distant relatives (like Rye or Agropyron) with non-homologous genomes. These are the most challenging, requiring advanced chromosome engineering to succeed.
2. The Hierarchy of Genetic Resources: Understanding Gene Pools
Successful molecular breeding requires a precise understanding of genomic homology. The three-tier gene pool system dictates the technical approach required for trait transfer, ranging from simple hybridization to complex cytogenetic manipulation of the Ph1 locus.
The “Security Guard”: Overcoming the Ph1 Barrier
Why isn’t this “copy and paste” process easier? Because the wheat genome has a sophisticated “security guard” known as the Ph1 (Pairing homoeologous) locus. This allele, located on chromosome 5B, strictly ensures that wheat chromosomes only pair with their exact matches. This protects the plant’s identity as a hexaploid species, but it effectively blocks “alien” chromosomes from swapping information with wheat.
To bypass this guard, we use specialized biological “skeleton keys”:
- The CS ph1b Mutant: A Chinese Spring (CS) stock carrying a small intercalary deficiency at the Ph1 locus. The absence of this suppressor promotes homoeologous recombination, allowing for the precise exchange of small alien segments into wheat chromosomes.
- Monosomic 5B Stocks: Utilizing wheat lines deficient in chromosome 5B as the female parent in hybridization. In the absence of 5B, meiotic pairing between wheat and alien chromosomes (e.g., from Ae. kotschyi or Ae. peregrina) is significantly induced.
- Pairing Suppressors: Utilizing species like Ae. speltoides, which naturally carry dominant suppressors of the Ph1 locus (such as Su1-Ph1 and Su2-Ph1), effectively unlock meiotic pairing in F1 hybrids.
- Radiation: Ionizing radiation can be used to physically break chromosomes, forcing a translocation (a swap) between species.
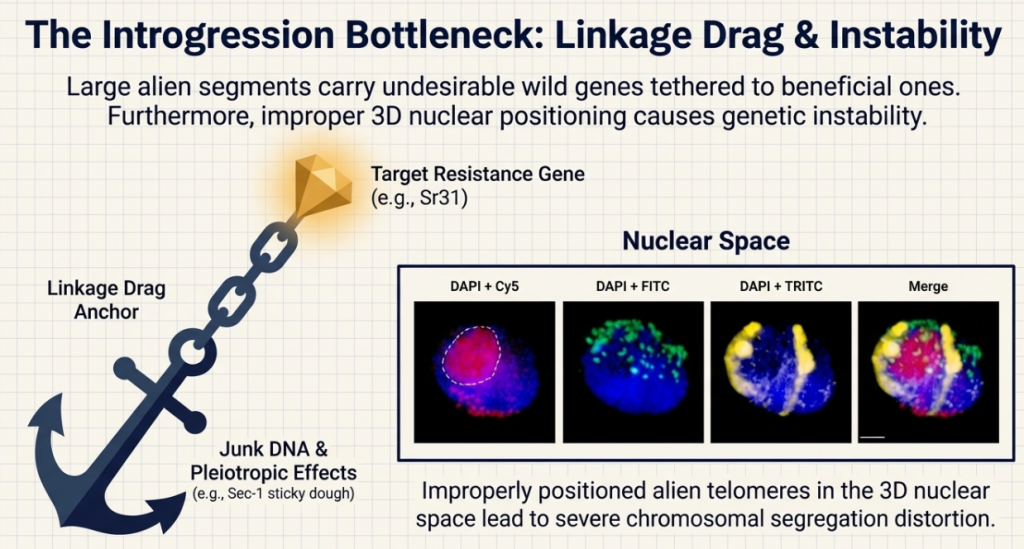
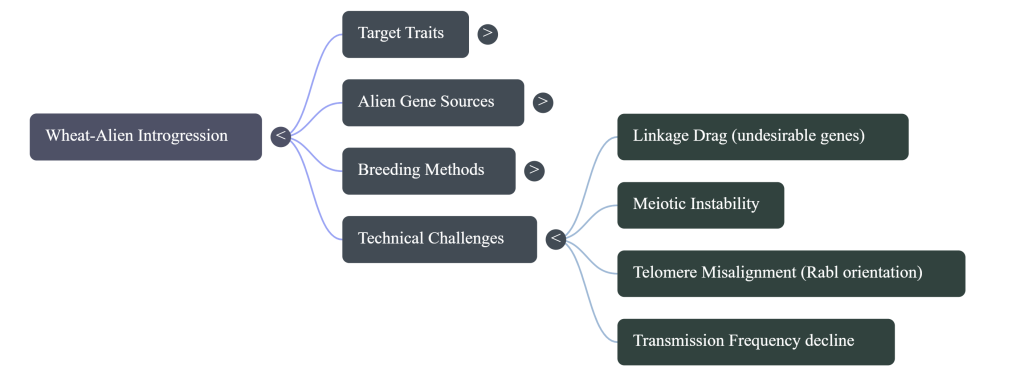
Technical Term: Linkage Drag This is the ultimate breeder’s challenge. Imagine you want to import a “shield” (a resistance gene) from a wild relative, but it comes attached to “baggage” (negative traits). In wheat, we might successfully transfer rust resistance, but accidentally bring along the genes for “sticky dough” or reduced yield. Solving this requires precision engineering to keep the shield while discarding the baggage.
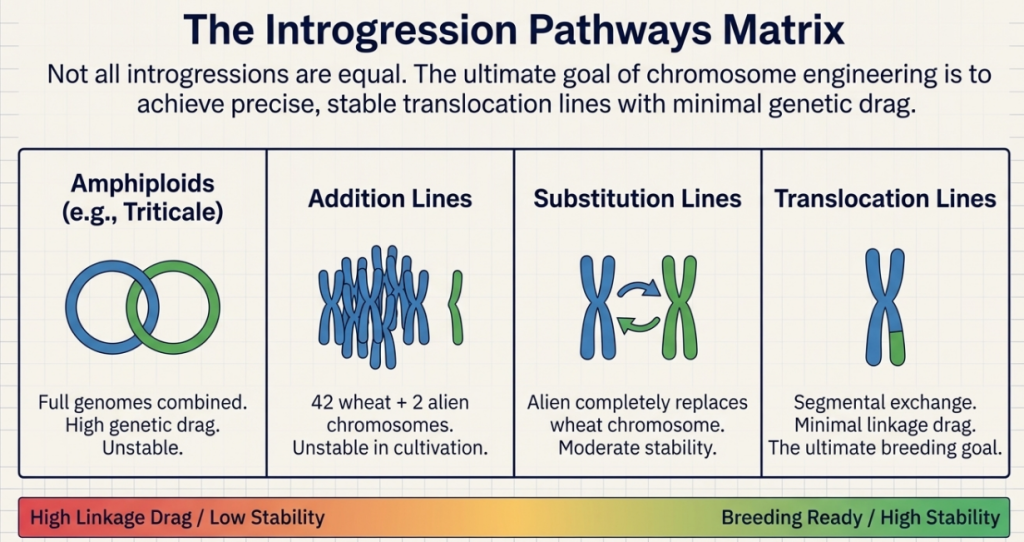
Technical Classification of Wheat Genetic Resources
| Gene Pool Category | Representative Species | Genomic Formula | Hybridization Potential / Barriers |
|---|---|---|---|
| Primary | Ae. tauschii, T. boeoticum, T. dicoccoides | DD, AbAb, AABB | Highly accessible, allowing direct hybridization and homologous recombination. T. dicoccoides and Ae. tauschii are the direct progenitors of the wheat A, B, and D genomes—minimal biological barriers. |
| Secondary | Ae. cylindrica, Ae. ventricosa, T. timopheevi | CCDD, DDNN, AAGG | Moderate accessibility; shares at least one genome with wheat. Requires focused breeding protocols but allows for relatively direct gene transfer via homoeologous pairing. |
| Tertiary | Secale cereale (Rye), Leymus racemosus, Thinopyrum junceiforme | RR (Rye), Perennial (Leymus) | Complex accessibility; non-homologous genomes. Requires precision chromosome engineering, irradiation, or Ph1 manipulation to overcome complete sterility and bypass linkage drag. |
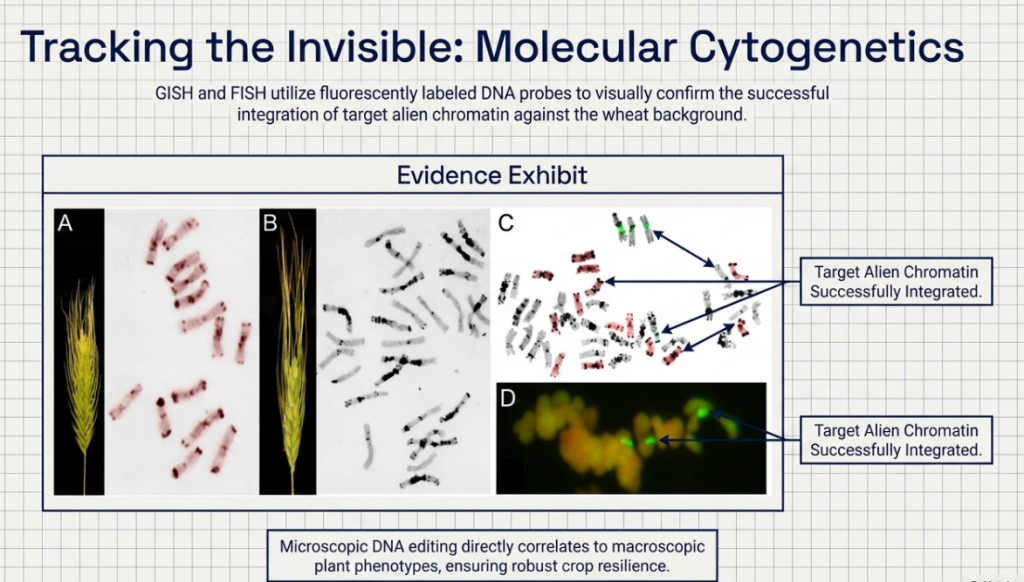
While the tertiary pool—specifically species like Leymus and Thinopyrum—is historically difficult to utilize, it represents the most significant frontier for novel variation. Transfers from this pool require specialized tools such as GISH (Genomic In Situ Hybridization) and FISH (Fluorescent In Situ Hybridization) to verify the stability and position of introgressed segments within the wheat nucleus.
The Three Pillars of Genetic Enhancement
Our research into these introgression lines has revealed three core benefits that will define the future of wheat:
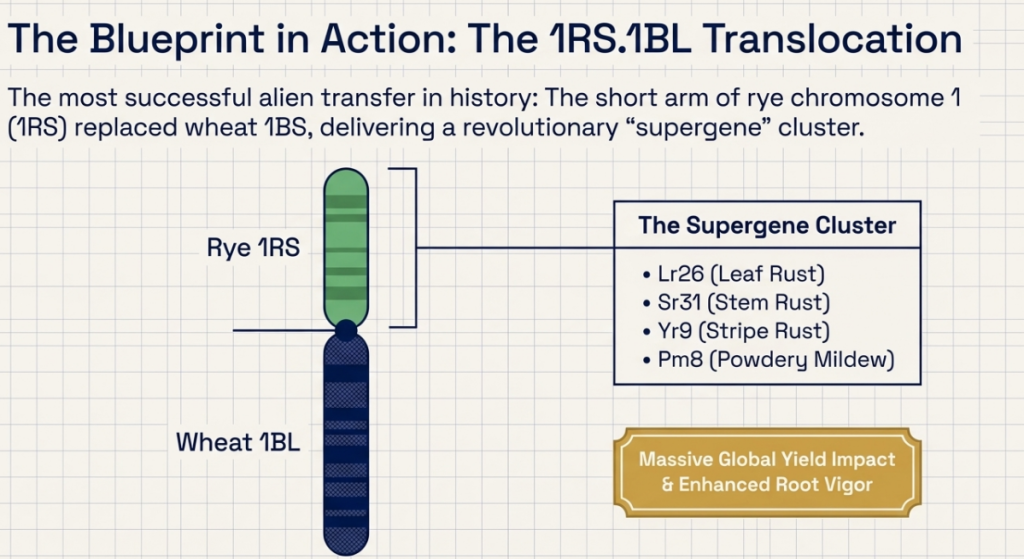
- Durable Resistance: We are using alien genes to build a “multi-layered shield.” For instance, rye chromosomes 4R, 5R, and 6R provide novel resistance against Stripe Rust, while 1R, 2R, and 6R protect against the Syrian Hessian fly and the Russian wheat aphid.
- Biofortification (The Micronutrient Power-Up): Beyond Iron and Zinc, the GPC-B1 allele (from wild emmer) provides a 29% increase in Manganese (Mn). Most importantly, the 1R(1D) substitution lines and Leymus introgressions allow us to achieve a “golden ratio”: high Fe and Zn density while simultaneously lowering toxic Cadmium (Cd) levels.
- End-Use Quality: While wild genes can sometimes cause “baggage” like sticky dough, certain introgressions from Leymus actually improve gluten strength, ensuring that more nutritious wheat can still produce the high-quality bread consumers demand.
Characterization of Alien Genetic Donors and Chromosomal Architecture
Taxonomically, alien genetic donors represent a broad spectrum within the tribe Triticeae. The primary cytogenetic challenge in their utilization is the introgression of non-homologous chromatin into the wheat genome. Because these species often lack natural pairing affinity with wheat, specialized chromosome engineering is required to facilitate stable transfers and mitigate the disruption of genomic integrity.
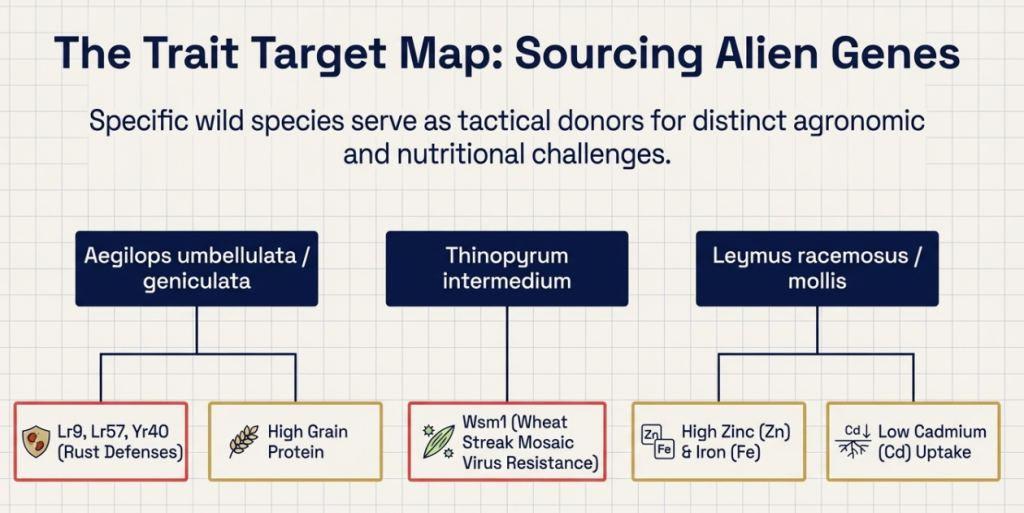
The following table summarizes the key alien donors and the specific genomic contributions evaluated for modern breeding:
Key Alien Genetic Donors and Genomic Contributions
| Species | Genome Formula | Target Traits Identified |
|---|---|---|
| Secale cereale (Rye) | RR | Rust resistance (Sr59, Yr83), Hessian fly/RWA resistance, Allelopathy |
| Leymus racemosus | JjNs / LrLr | High Zn and Fe, Fusarium head blight resistance (Fhb3) |
| Leymus mollis | JjNs / LmMm | High Zinc, Russian Wheat Aphid resistance, low Cadmium uptake |
| Thinopyrum spp. (T. junceiforme, T. intermedium) | JJ, Jj, St | Stem rust resistance, Powdery mildew resistance |
Breeding strategies are dictated by the accessibility of these gene pools. The primary gene pool (GP-1) consists of species sharing homologous genomes with wheat, allowing for direct recombination. The secondary gene pool (GP-2), including species like Aegilops speltoides, shares at least one genome in common. The tertiary gene pool (GP-3), which includes Agropyron and Thinopyrum, possesses non-homologous genomes (“St”, “J”, “E”), requiring advanced cytogenetic manipulations to induce homeologous pairing and stabilize the resulting introgressions.
3. Methodological Framework: From Raw Germplasm to Adapted Lines
The breeding objective has shifted from crude wide hybridization toward precision chromosome engineering. The strategic goal is the development of stable, “compensatory” translocations where alien segments replace their homoeologous wheat counterparts without compromising agronomic performance.
The Precision Crossing Pipeline
- Initial Wide Hybridization: Crossing wheat (female) with alien donors to generate amphiploids. Success at this stage relies on high-crossability cultivars like Fukuhokomugi or Chinese Spring (carrying kr1, kr2, kr3, kr4 alleles) and the deployment of embryo culture to bypass post-fertilization barriers.
- Translocation Induction:
- Spontaneous Translocations: Utilizing centromeric breakage and fusion (Robertsonian translocations) during meiosis. While easier to manage, these often result in large introgressions.
- Radiation-Induced Translocations: Utilizing ionizing radiation to break chromosomes. Critically, pollen irradiation is often preferred over seed irradiation—particularly for transferring Fe/Zn traits from species like Ae. kotschyi—as it facilitates higher seed set and more targeted transfers (Sharma et al. 2018b).
- Stabilization and Backcrossing: Repeated backcrossing with elite adapted varieties to restore fertility and eliminate deleterious “baggage,” moving toward disomic substitution or addition lines.
The methodology must differentiate between compensatory translocations (stable and commercially viable) and non-compensatory ones, which often suffer from extreme linkage drag and quality degradation.
4. Mitigating Linkage Drag and Protecting End-Use Quality
The integration of alien germplasm presents significant economic risks. “Linkage drag” occurs when a target gene is introgressed alongside deleterious alleles. A classic example is the Sec-1 locus from rye, which, in 1RS translocations, replaces the wheat Glu-B3/Gli-B1 locus. This introduces rye secalins, leading to the “sticky dough” issue that significantly penalizes commercial bakers.
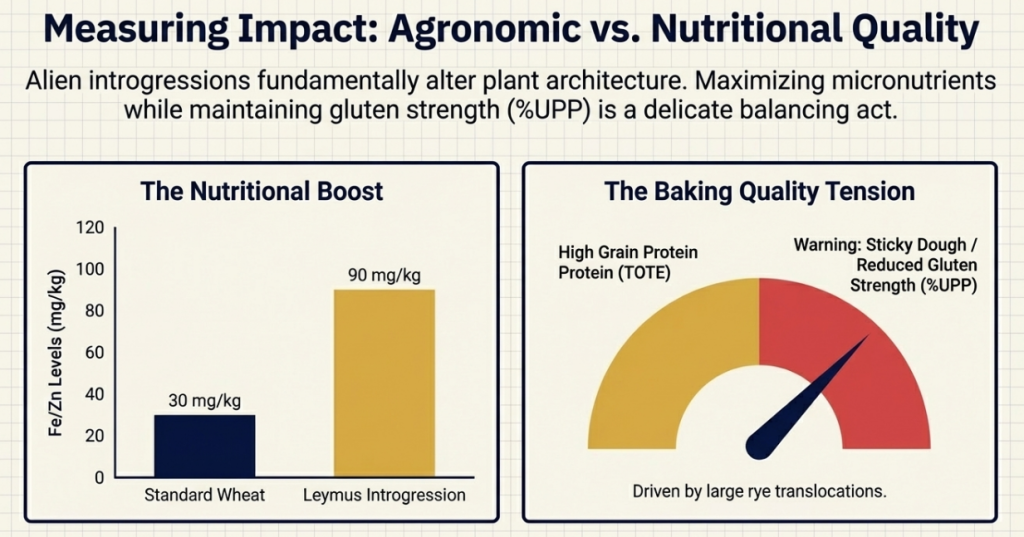
Quality Metrics and Biofortification Strategy
To maintain market competitiveness, breeders must monitor specific indicators:
Strategic ” Prizes”: Introgressions from Leymus racemosus and Thinopyrum junceiforme offer high Fe/Zn concentrations (ranging from 53.7 to 83.4 mg/kg for Zn) while maintaining low Cadmium (Cd) levels—addressing both the “Hidden Hunger” market and food safety standards.
Gluten Strength: Measuring the percentage of unextractable polymeric protein (%UPP) and total extractable protein (TOTE). For instance, stacking Glu-B1 (7+8) or Glu-D1 (5+10) subunits can help overcome the quality defects introduced by secalins.
Nutritional Density: The GPC-B1 (NAM-B1) locus on chromosome 6BS, derived from T. dicoccoides, is the cornerstone of biofortification. This NAC transcription factor accelerates senescence and increases nitrogen, Fe, and Zn remobilization.
5. Evaluation of Durable Disease Resistance: Rusts and Powdery Mildew
The biological threat posed by Puccinia graminis tritici (stem rust) and Puccinia striiformis tritici (stripe rust) remains a paramount concern for global wheat productivity. The emergence of the TTKSK (Ug99) race lineage has rendered many traditional resistance genes obsolete, necessitating the isolation of novel, durable alleles.
A significant advancement in this area is the characterization of the Sr59 gene. Originating from the 2RL chromosome of rye, Sr59 was developed specifically through a 2DS.2RL Robertsonian translocation utilizing the ph1b mutant to facilitate the transfer. In rigorous seedling and adult plant assays, Sr59 demonstrated high-level resistance to TTKSK, providing a critical alternative to the Sr31 gene, which has faced recent breakdowns in various global wheat-growing regions.
Regarding stripe rust, multiple introgression lines exhibited robust resistance to 12 highly virulent European and Asian races. High-performing chromosomal configurations include:
- 4R, 5R, and 6R substitutions: These lines, replacing the 4D, 5D, and 6D wheat chromosomes respectively, demonstrated resistance across nearly all tested isolates.
Furthermore, field evaluations indicated that rye chromosomes 1R, 2R, 3R, and 5R are strongly correlated with reduced infection levels of Powdery Mildew and Zymoseptoria tritici. These introgressions offer a foundational multi-pathogen defense strategy for developing resilient cultivars.
6. Resistance Profiling for Syrian Hessian Fly (HF) and Russian Wheat Aphid (RWA)
Economic losses attributed to the Syrian Hessian fly and the Russian wheat aphid (RWA) underscore the necessity of host resistance as the most sustainable management strategy.
Assessment of winter wheat alien introgressions revealed several accessions with 100% resistance to the Syrian Hessian fly. Notable configurations include:
- Kr 08-59 and Kr 08-60: Carrying 1R.1D and 1R.1D + 6R.6D configurations.
- Kr 08-91, Kr 08-94, and Kr 08-95: Utilizing T2RL.2BS translocations.
From a cytogenetic monitoring perspective, the red coleoptile color serves as a vital morphological marker for the 2BS.2RL translocation, allowing for rapid identification of resistant material in breeding populations. For RWA, high-level resistance was synthesized from 3DL.3RS and 5AL.5RS translocations, alongside novel introgressions from Leymus mollis. These assets significantly widen the genetic base beyond the increasingly vulnerable H21 and H25 genes.
7. Nutritional Biofortification: Grain Mineral Density and Cadmium Mitigation
Addressing “hidden hunger”—the deficiency of essential micronutrients such as Iron (Fe) and Zinc (Zn)—requires the strategic biofortification of staple crops. Alien introgressions, particularly from the Leymus genus, exhibit transgressive segregation for mineral density compared to standard wheat.
Grain Mineral Concentration and Heavy Metal Mitigation
| Material Type | Iron (Fe) [Mean (Range)] mg/kg | Zinc (Zn) [Mean (Range)] mg/kg | Cadmium (Cd) [Range] mg/kg |
|---|---|---|---|
| Standard Wheat (n=5) | 45.0 (31.0–60.6) | 39.5 (34.5–48.7) | 0.07–0.12 (High) |
| Rye Donor (n=2) | 39.7 (38.1–41.2) | 35.2 (33.8–36.6) | 0.00 |
| Leymus spp. Donor (n=3) | 49.4 (41.4–59.1) | 75.8 (62.4–83.4) | 0.00–0.02 |
| Wheat–Rye Introgressions | 38.9 (22.7–64.2) | 54.8 (32.9–89.3) | 0.00–0.10 (Medium) |
| Wheat–Leymus Introgressions | 47.5 (43.0–51.9) | 63.6 (53.1–69.1) | 0.02–0.06 (Low) |
The success of Leymus racemosus and Leymus mollis in Zinc enhancement is particularly noteworthy. Nine specific lines were identified combining high Fe/Zn density with low Cadmium (0.02–0.07 mg/kg) concentrations. This nutritional improvement is biologically facilitated by the GPC-B1 locus (a NAC transcription factor), which accelerates senescence and optimizes the remobilization of minerals from the vegetative tissues to the developing grain.
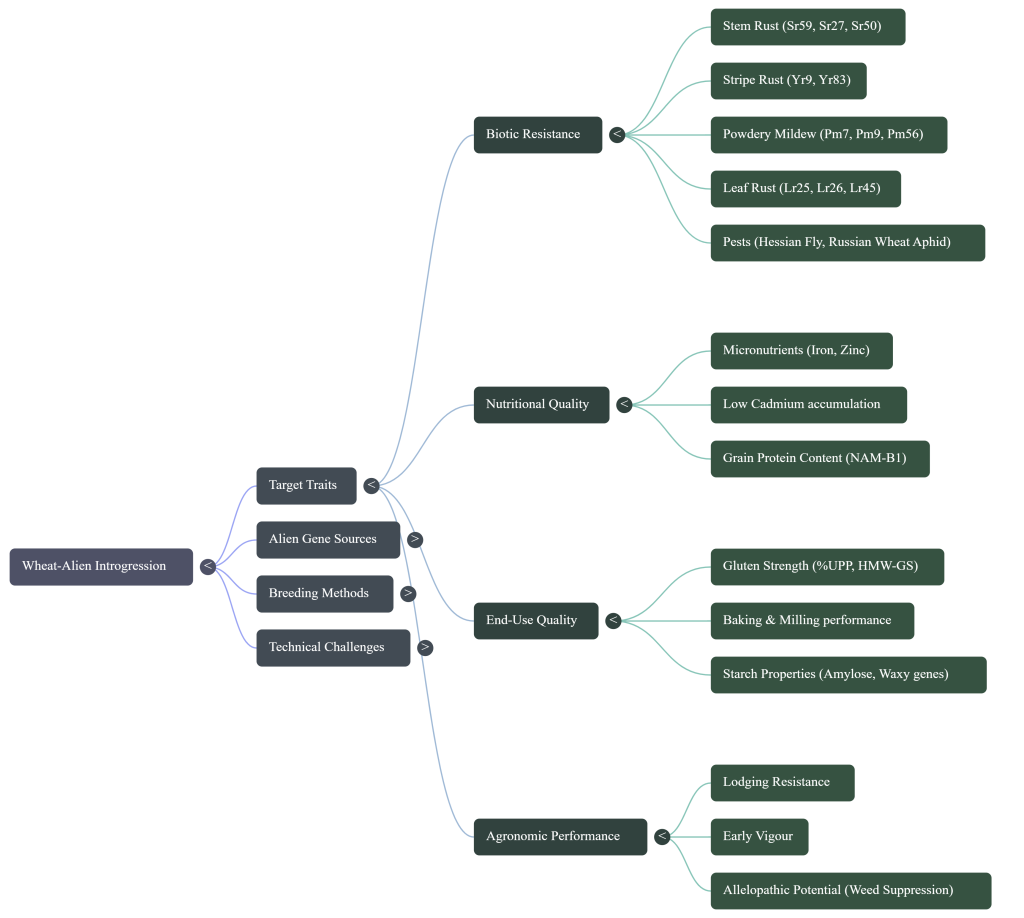
Summary of Strategic Trait Targets
The complexity of ph1b-mediated introgression is justified by the acquisition of elite genetic “prizes” identified in the source materials.
High-Value Trait Catalog
- Durable Resistance:
- Sr59: A novel gene from a 2DS.2RL translocation effective against all known stem rust races, including the TTKSK (Ug99) lineage.
- Yr83: Located on 6RL, providing durable stripe rust resistance.
- Pm21 and Pm56: Providing robust powdery mildew resistance.
- Pest Resistance:
- H21 (2R) and H25 (6R) for Syrian Hessian Fly resistance.
- Dn7 (1R) and L. mollis introgressions for Russian Wheat Aphid resistance.
- Biofortified Nutrition: The Gpc-B1 locus for enhanced protein, Fe, and Zn density.
- Agronomic Resilience: Allelopathic potential from rye (1R/2R) for weed suppression and 2BS.2RL translocations for improved lodging resistance.
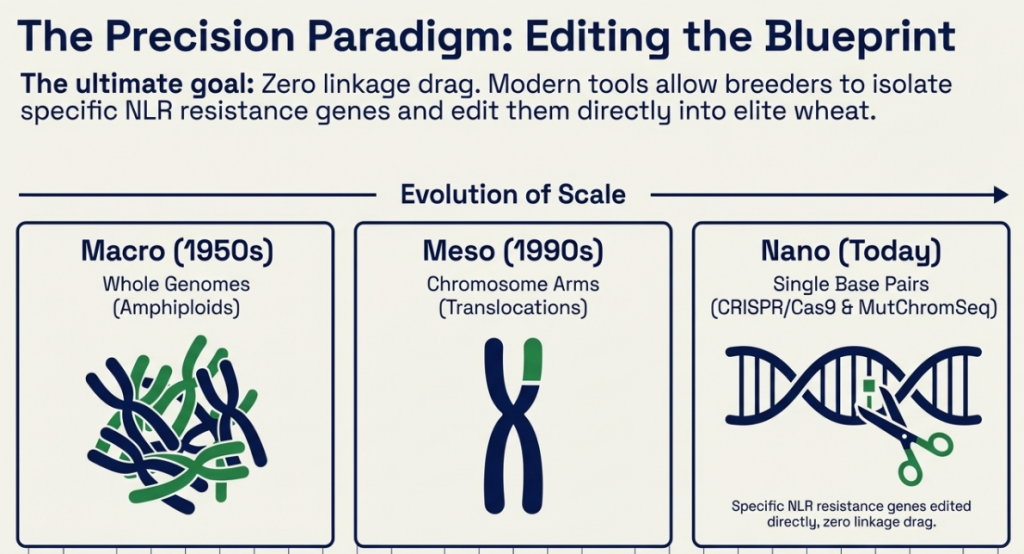
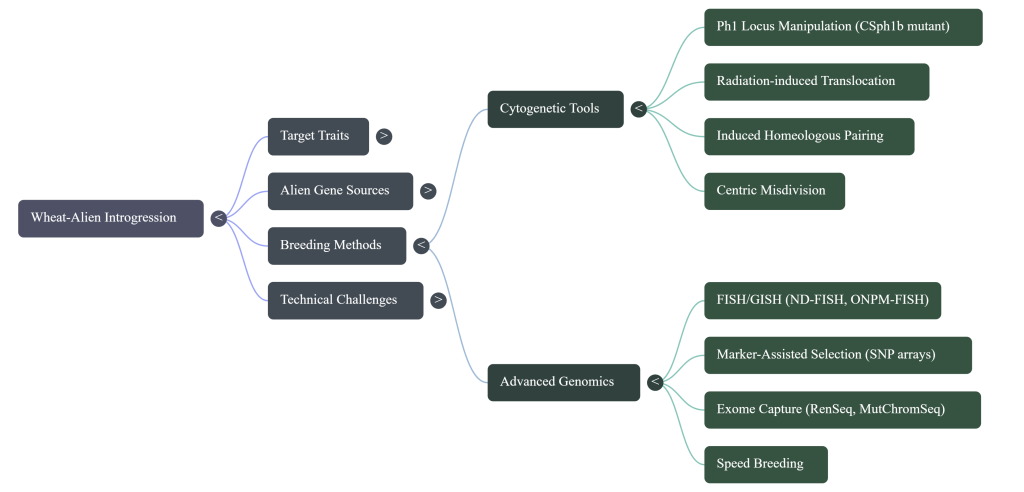
8. Advanced Breeding Methodologies and Chromosome Engineering: Overcoming Biological Barriers
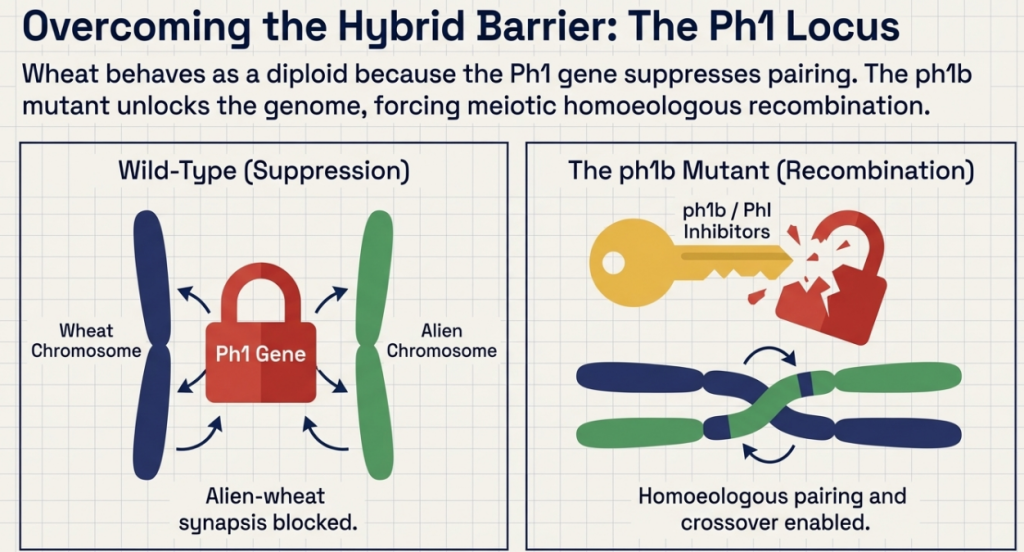
The primary hurdle in alien gene transfer is the Ph1 (Pairing homoeologous) locus on chromosome 5B, which restricts meiotic pairing to true homologs. Precision breeding requires bypassing this control to minimize linkage drag—the co-inheritance of deleterious wild traits.
1. Induced Homeologous Pairing via ph1b Mutants: Utilizing the ph1b mutant (a small intercalary deficiency in the Ph1 locus) is the gold standard. This allows for precise recombination between alien segments and wheat chromosomes, significantly reducing the size of the introgressed segment.
- CS ph1b Mutant Stocks: These allow for the induction of homeologous recombination, enabling the transfer of smaller, more refined alien segments rather than whole arms.
2. Radiation-Induced Translocations: Utilizing ionizing radiation (pollen or seed irradiation) to break chromosomes and force the fusion of alien segments, such as the transfer of HMWGS from Aegilops kotschyi.
- Pollen/Seed Irradiation: A critical tool for inducing translocations between non-homologous chromosomes when natural pairing is absent.
3. Spontaneous Translocations: Exploiting centromeric breakage and fusion in univalents during meiotic metaphase in wide hybrids (e.g., the 1RS.1BL translocation).
4. Substitution and Addition Lines: Systematically replacing wheat pairs (e.g., 1R substituting for 1D) or adding alien pairs to develop the foundational material for further segment reduction.
5. MutChromSeq and RenSeq: These technologies utilize mutational genomics and sequence capture to bypass traditional mapping, which often takes years. They allow for the rapid isolation and cloning of resistance genes (e.g., Sr22, Sr45) directly from complex genomes.
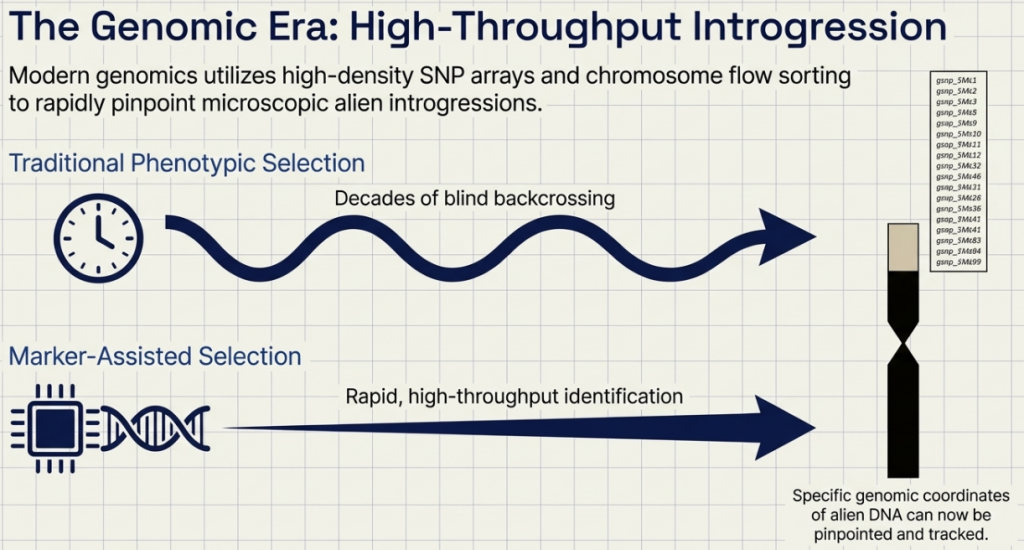
6. 90K SNP Arrays: High-throughput genotyping allows breeders to track alien chromatin with high resolution, ensuring the retention of target traits while eliminating unwanted donor DNA.
These tools collectively compress the breeding cycle, allowing for the rapid deployment of wild-derived traits into elite backgrounds.
Modern Precision Tools: To transition from random introgression to precision engineering, breeders now integrate Speed Breeding to accelerate generations, MutChromSeq for rapid gene cloning, and Genotyping-by-Sequencing (GBS) to map small alien segments with high resolution.
Baking Functionality Matrix: Alien Introgression Effects
| Introgression Type | Gluten Strength (%UPP) | Dough Functionality | Starch / End-Use Effect |
|---|---|---|---|
| 1BL.1RS (Rye) | Variable | High frequency of sticky dough due to secalins (Sec-1 locus). | Often used for yield but requires quality compensation. |
| 2BS.2RL (Rye) | Stable | Minor/manageable effects; no significant difference in alpha-amylase. | Preferred for combining multiple resistance with quality. |
| Leymus spp. | High | Increases unextractable polymeric protein (UPP). | Improves gluten strength and mineral density. |
| 1R+2R / 1R+4R | High | Significant increase in glutenin polymerization. | Synergistic effect on dough strength and protein content. |
9. Conclusion: The Future of Alien Breeding in Modern Wheat
To meet the 2050 global food demand, we must aggressively broaden the genetic base of Triticum aestivum. The strategic integration of the Triticeae gene pool is the only viable path to developing “climate-smart” wheat.
The Future Toolkit: CSI: Crop Science
We are no longer wandering in the dark as breeders did in the 1980s. Today, we have a “CSI: Crop Science” suite that makes alien introgression faster and more precise than ever before:
- RenSeq (The Genetic Search Engine): This tool allows us to scan the vast haystack of a wild genome to find the one specific “needle”—the resistance gene we need.
- SNP Arrays (9K, 90K): High-speed genotyping that acts like a GPS, telling us exactly where an alien segment has landed in the wheat genome.
- Speed Breeding (The Time Machine): By using controlled light and temperature, we can “fast-forward” the plant’s life cycle, achieving up to six generations a year. What used to take a decade now takes a fraction of that time.
Genomic Tool Inventory
| Tool Category | Specific Technology | Strategic Application |
|---|---|---|
| Cloning/Enrichment | RenSeq / MutChromSeq | Rapid isolation and cloning of resistance genes; notably utilized for cloning Sr22, Sr45, and Pm2. |
| Genotyping | 90K SNP Arrays / GBS | Precise mapping of alien segments and identification of SNP polymorphisms across genomes. |
| High-Throughput | Exome Capture Sequencing | Uncovering hidden variation and accelerating the identification of nucleotide-binding leucine-rich repeat (NLR) genes. |
| Breeding Acceleration | Speed Breeding | Drastic generation time reduction, allowing for multiple breeding cycles per year to rapidly stabilize introgressed lines. |

The focus of modern breeding has shifted from broad, unstable introgressions to precision engineering. By leveraging foundational genetic resources—specifically Secale cereale, Leymus spp., and Aegilops—and utilizing tools like ph1b manipulation and GISH/FISH identification, we can deploy genes like Sr59 and GPC-B1 with surgical accuracy. This ensures that the next generation of wheat cultivars offers not only high yield but also durable resistance and superior nutritional value.
Image Summary
Questions/Answers
1. How do rye chromosomes improve wheat’s resistance and nutrition?
Rye (Secale cereale) chromosomes and chromosome segments, particularly from the 1R, 2R, and 3R genomes, are among the most widely utilized alien genetic resources for improving bread wheat. These introgressions enhance wheat by providing robust resistance to a wide range of diseases and pests, increasing tolerance to environmental stresses, and boosting nutritional content.
Biotic Resistance: Diseases and Pests
Rye introgressions are a primary source of broad-spectrum disease resistance in modern wheat cultivars:
- Fungal Diseases: The most successful transfer is the 1RS chromosome arm (typically as 1BL.1RS translocations), which carries a “supergene” block of resistance genes: Sr31 (stem rust), Yr9 (stripe rust), Lr26 (leaf rust), and Pm8 (powdery mildew). Other rye chromosomes contribute resistance to Septoria tritici and Fusarium head blight. Novel resistance genes like Sr59 (stem rust) and Yr83 (stripe rust) have also been identified on rye chromosomes 2R and 6R.
- Pest Resistance: Rye chromosomes provide essential defense against destructive insects. Chromosome 1R carries genes for resistance to the Russian wheat aphid (Dn7) and cereal aphids (Gb2, Gb6). Chromosomes 1R, 2R, and 6R have also been found responsible for resistance to the Syrian Hessian fly (including genes H21 and H25).
- Allelopathic Potential: Rye is an excellent source of allelopathic activity, which allows wheat to better suppress weed growth, a trait particularly valuable for organic farming.
Abiotic Stress Tolerance
Rye chromatin significantly improves wheat’s resilience to harsh environmental conditions:
- Drought Tolerance: Introgressions from 1R, 1RS, and 3R are associated with robust and deeper root systems, increasing root biomass and allowing wheat to access water in deep soil layers during surface drying. This leads to greater yield stability under drought conditions.
- Soil Adaptation: Chromosome 3R provides tolerance to aluminum toxicity, a major constraint in acidic soils. Rye substitutions like 1R(1B) also help maintain high yield potential in soils with low fertility.
- Reproductive Vigor: Addition of rye chromosome 4R has been shown to increase anther length and pollen grain number, which can improve pollination efficiency.
Nutritional and Quality Enhancement
Introgressing rye chromosomes can diversify the nutritional profile of wheat grain:
- Micronutrients: Wheat lines with 1R, 2R, 3R, and 5R introgressions often exhibit significantly higher concentrations of Zinc (Zn) and Iron (Fe). Some of these lines also show a desirable combination of high micronutrients and low Cadmium (Cd) concentration.
- Protein and Fiber: Chromosomes 1R, 2R, 4R, and 6R are linked to increased grain protein concentration. Additionally, rye introgressions can enhance the content of health-promoting dietary fibers, such as arabinoxylan and beta-glucan.
Trade-offs in End-Use Quality
While rye introgressions provide many benefits, they often introduce linkage drag that negatively impacts bread-making quality. Specifically, the Sec-1 locus on the 1RS arm encodes secalins (γ-secalin and ω-secalin), which can cause weak and sticky dough. However, recent studies indicate that introgressions from other rye chromosomes, such as 2RL or 3R, may have fewer negative effects or even contribute positively to baking quality stability.
2. Which wild relatives offer the best traits for baking quality?
Wild relatives of wheat are considered invaluable reservoirs for enhancing baking and bread-making quality, primarily by introducing novel alleles for seed storage proteins that have been lost through the narrow genetic bottleneck of wheat domestication.
The most significant traits for baking quality are governed by high-molecular-weight glutenin subunits (HMW-GS) and low-molecular-weight glutenin subunits (LMW-GS), which are primarily located on the group 1 and group 6 chromosomes. Several wild relatives offer superior variants of these proteins:
Top Aegilops (Goatgrass) Species
- Aegilops geniculata (Mg and Ug genomes): This species is one of the most promising donors for baking quality. Chromosome 1Mg has been shown to positively affect grain hardness, protein content, gluten content, and sedimentation value. Additionally, 1Ug introgressions improve the rheological properties of dough by enhancing the microstructure of gluten.
- Aegilops longissima (Sl genome): Chromosome 1Sl is recognized for carrying genes that significantly enhance bread-making quality. Substitution lines containing 1Sl have demonstrated improved farinograph properties and bread-making scores compared to standard wheat.
- Aegilops umbellulata (U genome): This relative provides LMW-GS from the 1U genome that confer superior dough rheological properties and improve general bread-making quality. While some 1U fragments were initially reported to have negative effects on dough strength, specific LMW-GS substitution lines (Glu U3a and Glu U3b) have been shown to increase mixing properties and loaf volume.
- Aegilops comosa (M genome): Chromosome 1M has been reported to have a positive effect on seed morphology and bread-making quality traits.
- Aegilops biuncialis (Ub and Mb genomes): The 1Ub addition line carries HMW-GS associated with high baking quality, and 1Mb(1B) substitution lines have also shown favorable quality-associated glutenin subunits.
Other Key Wild Relatives
- Dasypyrum villosum (Hv or V genome): Introgressing chromosome segments from 1V has shown positive effects on the bread-making quality of common wheat by introducing superior seed storage protein genes.
- Wild Emmer (Triticum dicoccoides): While often used for yield components, it is the donor of the GPC-B1 (NAM-B1) gene. This gene increases grain protein content and enhances the remobilization of nutrients to the grain, which indirectly supports baking quality by increasing the total protein available for gluten formation.
- Rye (Secale cereale): While the common 1RS.1BL translocation is widely used for yield and resistance, it is notorious for decreasing baking quality by causing sticky dough and weak gluten due to the loss of wheat glutenin subunits and the addition of rye secalins. However, introgressions of the 3R chromosome have been shown to contribute positively to baking quality stability, especially under drought conditions.
- Agropyron elongatum (Tall wheatgrass): This relative possesses HMW-glutenins with the potential to improve end-product quality through targeted chromosome introgression.
Summary of Quality Improvements
| Wild Relative | Chromosome/Gene | Phenotypic Effect on Quality |
|---|---|---|
| Ae. geniculata | 1Mg, 1Ug | Increased grain hardness, gluten strength, and dough rheology. |
| Ae. longissima | 1Sl | Enhanced bread-making quality and farinograph properties. |
| Ae. umbellulata | 1U (LMW-GS) | Improved mixing properties, dough development time, and loaf size. |
| Ae. searsii | 1Ss (HMW-GS) | Novel pair of HMW-GS that improves quality in hexaploid wheat. |
| D. villosum | 1V | Positive effect on bread-making quality. |
| T. dicoccoides | GPC-B1 | Significantly higher grain protein and mineral content. |
3. Tell me more about the Sr59 stem rust resistance gene.
Sr59 is a novel and highly effective stem rust resistance gene that was recently introgressed into bread wheat from the long arm of rye chromosome 2R (2RL). It is considered one of the most valuable alien genetic resources for wheat improvement because it provides broad-spectrum resistance to all currently identified races of stem rust, including the highly virulent Ug99 (TTKSK) race lineage.
Discovery and Development
- Origin: The gene was originally identified in a Swedish winter wheat-rye disomic substitution line designated SLU238, where rye chromosome 2R replaced wheat chromosome 2D.
- Introgression Strategy: To utilize the gene in commercial wheat breeding while minimizing negative “linkage drag,” researchers transferred it into wheat through a new 2DS·2RL Robertsonian translocation.
- Chromosome Engineering: This translocation was achieved by crossing the SLU238 substitution line with the Chinese Spring ph1b mutant. The ph1b mutation disables the wheat Ph1 gene, which normally prevents recombination between wheat and alien chromosomes, thereby facilitating the transfer of the 2RL segment carrying Sr59 into the wheat genome.
Significance to Wheat Breeding
- First of its Kind: Prior to the discovery of Sr59, no stem rust resistance genes had ever been reported on rye chromosome 2R. Previous rye-derived stem rust genes, such as the widely deployed Sr31, originated from chromosome 1R.
- Durability: While Sr31 provided durable resistance for over 30 years, it has recently been overcome by new stem rust races in Africa. In contrast, Sr59 remains effective against these new virulent races, making it a critical tool for global food security.
- Breeding Tools: Modern breeding efforts to integrate Sr59 into adapted wheat varieties utilize advanced technologies such as genotyping-by-sequencing (GBS) and DNA marker-assisted selection to ensure the gene is successfully transferred with minimal non-target alien chromatin.
Beyond stem rust resistance, other lines carrying the 2RL chromosome arm have demonstrated good agronomic performance and multiple resistance to several races of powdery mildew, though these traits are distinct from the specific resistance conferred by the Sr59 gene.
4. How do 1RS translocations cause weak and sticky dough?
The weak and sticky dough performance associated with 1RS translocations (most commonly the 1BL.1RS translocation) is primarily caused by a “double effect” of genetic loss and alien gene introduction: the deletion of essential wheat glutenin proteins and the introduction of rye secalin proteins.
1. Deletion of Critical Wheat Proteins
When the 1RS rye chromosome arm replaces the 1BS wheat arm, several loci critical for dough strength are lost:
- Loss of Glu-B3 and Gli-B1: The deletion of the short arm of wheat chromosome 1B results in the loss of these loci, which encode important low-molecular-weight (LMW) glutenin subunits and gliadins.
- Weak Dough: Because these glutenin subunits are vital for the elasticity and structural integrity of the gluten network, their absence leads to significantly reduced dough strength, or “weak dough“.
2. Introduction of Rye Secalins
The introgressed 1RS arm brings with it rye-specific storage protein genes that interfere with wheat’s standard processing properties:
- The Sec-1 Locus: This locus on the 1RS arm encodes high-molecular-weight proteins known as γ-secalin and ω-secalin.
- Sticky Dough: The presence of these secalins is the primary factor responsible for increased dough stickiness. This stickiness makes the dough difficult to handle during industrial processing and baking.
3. Impact on Gluten Polymerization
The exchange of wheat high-molecular-weight (HMW) glutenin subunits for rye secalins disrupts the ability of gluten proteins to polymerize effectively during dough formation. This polymerization is a major determinant of the viscoelastic properties required for high-quality bread-making.
Potential Mitigation
While these traits have traditionally limited the use of 1RS in premium baking wheats, recent research has shown that baking quality can be improved if gluten protein genes are restored in these 1BL.1RS translocation lines. Additionally, introgressions from other rye chromosomes, such as chromosome 3R, have been found to contribute positively to baking quality stability without causing the same stickiness issues associated with 1RS.
5. Explain the role of the Ph1 locus in alien breeding.
The Ph1 (Pairing homoeologous 1) locus is a key regulator located on the long arm of wheat chromosome 5B that ensures the stable, diploid-like meiotic behavior of polyploid wheat. Its primary role in alien breeding is as a genetic gatekeeper that must be manipulated to allow the transfer of genes from wild relatives into the wheat genome.
The Role and Mechanism of Ph1
- Enforcement of Homologous Pairing: In its normal (wild-type) state, the Ph1 gene restricts meiotic pairing and recombination strictly to homologous chromosomes (identical partners). It prevents pairing between homoeologous chromosomes (genetically related but non-identical partners from different subgenomes or alien species).
- The Barrier to Breeding: Because chromosomes from wheat’s wild relatives are homoeologous rather than homologous, the presence of an active Ph1 locus effectively blocks the genetic recombination necessary to integrate alien DNA into the wheat background.
Cytogenetic Manipulation for Gene Transfer
To facilitate the introgression of desirable traits, breeders must bypass the Ph1 system using several strategies:
- The ph1b Mutant: The most widely used tool is the ph1b mutant, which features a large deletion of the 5BL region containing the Ph1 gene. This mutation promotes meiotic synapsis and allows wheat chromosomes to pair and recombine with their alien homoeologous counterparts.
- External Suppressors: Pairing can also be induced by crossing wheat with specific accessions of Aegilops speltoides or Ae. mutica that carry dominant genes (such as Su1-Ph1 or Su2-Ph1) capable of inhibiting the activity of the wheat Ph1 locus.
- Nullisomic Lines: Using wheat lines that completely lack chromosome 5B (nullisomics) also removes the pairing restriction, though this can lead to greater genomic instability.
Benefits in Alien Breeding
Manipulating the Ph1 locus is essential for “chromosome engineering” to achieve high-precision gene transfer:
- Reduction of Linkage Drag: Targeted homoeologous recombination allows for the transfer of small alien chromosome segments rather than entire chromosomes. This is critical for minimizing “linkage drag,” where undesirable wild traits (e.g., poor yield or baking quality) remain linked to the beneficial target gene.
- Genetic Stability: While alien addition or substitution lines are often unstable, translocations produced through Ph1 manipulation are more likely to be genetically stable and breeder-friendly.
Post-Recombination Restoration
Once the desired alien segment has been successfully translocated into the wheat genome, the ph1b mutant chromosome must be eliminated. By backcrossing the recombinant lines with wild-type wheat, the functional Ph1 locus is restored, ensuring that the newly modified genome pairs normally and remains stable in future generations.
6. How does Aegilops geniculata specifically affect grain hardness and gluten?
Introgressions from Aegilops geniculata (an allotetraploid wild relative with the Mg and Ug genomes) specifically enhance wheat quality by improving grain texture and the functional properties of gluten proteins. These effects are primarily attributed to chromosomes in homoeologous group 1.
Impact on Grain Hardness
Chromosome 1Mg is identified as a major contributor to grain texture. Research has demonstrated that its presence in wheat lines results in:
- Increased Grain Hardness: The 1Mg chromosome positively affects grain hardness and volume-weight.
- Improved Milling/Baking Parameters: In addition to hardness, 1Mg introgressions are associated with higher Zeleny sedimentation values, which is a key indicator of baking strength.
Impact on Gluten and Dough Properties
Aegilops geniculata provides novel variants of seed storage proteins that improve the functional “strength” of the dough:
- Gluten Content and Strength: Chromosome 1Mg has a significant positive effect on total gluten content and protein concentration.
- Gluten Microstructure: Introgressions of chromosome 1Ug have been shown to improve the rheological properties of dough. This is achieved by altering the composition and microstructure of the gluten network, leading to better dough elasticity and stability.
- Baking Quality stability: These group 1 introgressions collectively contribute to superior baking quality, enhancing the performance of wheat flour in bread-making.
Nutritional and Protein Support
Beyond gluten strength, other chromosomes from Ae. geniculata contribute to the overall protein profile:
- Protein Content: Introgressions of chromosomes 2Ug, 4Ug, 5Ug, 7Ug, 2Mg, and 7Mg have all been reported to increase the total seed protein content in bread wheat.
- Dietary Fiber: Chromosomes 5Mg and 5Ug have also been noted for their positive effects on grain dietary fiber and beta-glucan content, further enhancing the nutritional value of the flour.
7. What makes Leymus species so effective for improving gluten strength?
Leymus species, specifically Leymus racemosus and Leymus mollis, are effective for improving gluten strength because they contribute novel major genes and quantitative trait loci (QTL) that enhance the functional properties of wheat gluten.
The effectiveness of Leymus introgressions is characterized by the following factors:
- High %UPP Values: Gluten strength in Leymus introgression lines is often measured as the percentage of unextractable polymeric protein (%UPP). High levels of %UPP are strongly correlated with superior gluten strength and bread-making quality, and lines carrying Leymus segments have been found to significantly exceed standard wheat cultivars in these values.
- Unique Genetic Resource: Unlike the widely used rye 1RS translocation, which often results in “linkage drag” that weakens dough and makes it sticky, certain Leymus introgressions can improve functional properties without these negative side effects. Leymus species possess a unique evolutionary history that provides genetic variation for seed storage proteins no longer present in the narrow genetic base of cultivated wheat.
- Matrix and Structural Building: The improved structural properties of proteins derived from Leymus make these lines particularly valuable for the development of novel food products, such as meat analogs. The high gluten strength allows for the formation of the fibrous, highly ordered network required to mimic meat texture.
- Dual-Benefit Traits: Introgressions from Leymus often provide a combination of enhanced nutrition and functional quality. For example, lines obtained from crosses with L. racemosus and L. mollis have demonstrated both high gluten strength and significantly higher concentrations of micronutrients like zinc.
Research into these species has identified specific chromosomes, such as 7Lr#1 from L. racemosus, that contribute valuable traits like resistance to Fusarium head blight alongside these quality improvements. Modern breeding tools, such as marker-assisted selection and GBS (Genotyping-by-sequencing), are now being used to transfer these beneficial Leymus segments into elite wheat backgrounds while minimizing undesirable alien chromatin.
8. How was the CS ph1b mutant used to transfer Sr59?
The Chinese Spring (CS) ph1b mutant was used to transfer the Sr59 stem rust resistance gene from rye into wheat by inducing homoeologous recombination, which allowed the alien gene to be integrated into a wheat chromosome while minimizing the amount of unwanted rye DNA.
According to the sources, the transfer process involved the following steps:
- Donor Selection: The Sr59 gene originates from the long arm of rye chromosome 2R (2RL). It was originally identified in a Swedish winter wheat-rye disomic substitution line designated SLU238, where the rye 2R chromosome replaced wheat chromosome 2D.
- The Crossing Strategy: To move the gene into a more useful wheat background, researchers crossed the SLU238 line with the Chinese Spring ph1b mutant.
- The Role of the ph1b Mutation: The Ph1 gene (located on wheat chromosome 5B) normally acts as a “gatekeeper,” strictly enforcing homologous pairing and preventing wheat chromosomes from recombining with their alien counterparts (homoeologues). The ph1b mutant has a large deletion of the 5BL region where this gene resides. By using this mutant, researchers disabled the pairing restriction, thereby promoting meiotic synapsis and recombination between the wheat and rye chromosomes.
- The Resulting Translocation: This cytogenetic manipulation facilitated a new 2DS·2RL Robertsonian translocation. This specific translocation successfully transferred the Sr59 gene into wheat on a stable chromosomal segment.
Importance of This Approach
Using the ph1b mutant is essential for chromosome engineering because it helps reduce linkage drag. Without this manipulation, breeders would often be forced to transfer entire alien chromosomes or large arms, which frequently carry deleterious traits that reduce yield or quality. The ph1b-mediated recombination allows for more precise transfers of smaller alien segments carrying target genes like Sr59 into adapted wheat varieties.
Once the Sr59 segment is successfully translocated, the ph1b mutant chromosome must be eliminated through further backcrossing with wild-type wheat to restabilize the genome for commercial use.
9. What other resistance genes come from rye chromosome 2R?
Beyond the recently discovered Sr59 gene, rye chromosome 2R is a well-documented source of genetic resistance against several significant wheat diseases and pests:
Named Resistance Genes
Several specific resistance genes have been identified and mapped to chromosome 2R:
- Lr25 and Lr45: These are leaf rust resistance genes originating from the 2R chromosome.
- Pm7: This gene provides resistance to powdery mildew.
- H21: This is a specialized gene for resistance to the Hessian fly (Mayetiola destructor).
Broad-Spectrum Resistance Traits
In addition to named genes, specific segments of chromosome 2R are associated with broader resistance profiles:
- Powdery Mildew: Lines carrying the long arm of rye chromosome 2R (2RL) have demonstrated multiple resistance against several different races of powdery mildew.
- Hessian Fly: Chromosome 2R is specifically responsible for resistance to the Syrian Hessian fly biotype.
- Allelopathic Potential: Rye chromosome 2R (along with 1R) has been identified as a primary source of allelopathic activity in wheat-rye introgression lines. This trait allows the plant to naturally suppress weed growth through root exudates, which is highly beneficial for sustainable and organic farming.
While many of these genes have seen limited use in commercial agricultural production to date compared to the 1RS “supergene” block, they remain critical resources for diversifying resistance in modern wheat cultivars.
10. How does chromosome 3R contribute to baking quality stability?
Rye chromosome 3R contributes to baking quality stability primarily by maintaining functional properties under environmental stress, particularly drought conditions. Unlike the widely used 1RS translocation, which often results in weak and sticky dough, 3R introgressions offer the following benefits:
- Environmental Stability: Research indicates that introgressions from the 3R chromosome contribute positively to baking quality stability across various drought treatments. This makes it a valuable resource for maintaining end-use consistency in changing climates.
- Gluten Strength: Studies have identified high gluten strength in alien wheat introgression lines carrying chromosome 3R. Gluten strength is the most critical criteria for baking quality, typically measured by instruments like the alveograph.
- Nutritional Support: Chromosome 3R is also associated with a higher grain protein concentration and increased levels of essential minerals. Specifically, lines with 3R introgressions have been found to contain high concentrations of Iron (Fe) and Zinc (Zn) while maintaining desirably low Cadmium (Cd) levels.
- Abiotic Stress Resilience: The chromosome provides traits that support overall plant health and yield stability, such as early drought tolerance and aluminum tolerance in acidic soils. These factors indirectly support quality by ensuring the plant can properly remobilize nutrients to the grain even under stress.
In summary, while other rye chromosomes like 1R provide high disease resistance but can compromise dough handling, chromosome 3R is distinguished by its ability to bolster gluten strength and protein concentration without those negative side effects, particularly under stressful growing conditions.
11. Which specific secalins are responsible for making dough sticky?
The specific secalins responsible for making dough sticky are γ-secalin and ω-secalin.
According to the sources, these proteins have the following characteristics and effects:
- Genetic Origin: These proteins are encoded by the Sec-1 locus, which is located on the short arm of rye chromosome 1R (1RS).
- Mechanism of Quality Reduction: In widely used wheat-rye translocations like T1BL·1RS, these rye storage proteins are introduced into the wheat genome at the same time that essential wheat proteins—specifically low-molecular-weight glutenin subunits and gliadins at the Glu-B3/Gli-B1 loci—are deleted.
- Resulting Texture: The loss of the wheat glutenin subunits weakens the dough, while the presence of the introduced rye secalins directly increases dough stickiness, making the flour difficult to handle and decreasing overall baking quality.
While these secalins negatively impact processing, the 1RS arm that carries them is often retained in breeding programs because it also provides a “supergene” block of resistance against major diseases like stem rust (Sr31), stripe rust (Yr9), and powdery mildew (Pm8).
12. How do breeders manipulate the Ph1 locus for gene transfer?
The Ph1 (Pairing homoeologous 1) locus, located on the long arm of wheat chromosome 5B, is the primary genetic regulator that enforces diploid-like behavior in polyploid wheat by restricting meiotic pairing to strictly homologous chromosomes. In its wild-type state, this gene prevents wheat chromosomes from synapsing and recombining with homoeologous (genetically related but non-identical) chromosomes from wild relatives, acting as a major barrier to alien gene transfer.
To bypass this barrier and facilitate the introgression of desirable alien traits, breeders employ several strategies to manipulate or suppress the Ph1 system:
1. Use of Recessive Mutants (ph1b and ph1c)
The most common method for inducing homoeologous recombination is the use of the ph1b mutant in hexaploid bread wheat.
- Mechanism: This mutant features a large radiation-induced deletion of the 5BL region where the Ph1 gene resides. The absence of the functional gene allows wheat chromosomes to pair and exchange genetic material with alien counterparts.
- Durum Wheat: A similar mutation, ph1c, has been developed for tetraploid durum wheat to achieve the same effect.
- Induced Rearrangements: This mutant has been successfully used to transfer various genes, such as the stem rust resistance gene Sr59 from rye and various resistance traits from Aegilops species.
2. Suppressors from Wild Relatives
Breeders can also inhibit the Ph1 locus by crossing wheat with specific accessions of wild relatives that carry dominant suppressor genes.
- Aegilops speltoides: This species is a primary source of such genes, carrying PhI, Su1-Ph1 (on chromosome 3S), and Su2-Ph1 (on chromosome 7S).
- Mechanism: When introduced into wheat-alien hybrids, these genes effectively “turn off” the wheat Ph1 system, promoting high levels of homoeologous pairing and recombination even in the presence of the 5B chromosome.
3. Chromosome 5B Deficiency and Removal
Directly removing or reducing the dosage of chromosome 5B also triggers homoeologous pairing:
- Nullisomic Lines: Using wheat lines that completely lack the 5B chromosome (nullisomic 5B) removes the pairing restriction entirely, though this often leads to meiotic instability.
- Monosomic 5B Lines: Crossing an alien relative with a wheat line that has only one copy of chromosome 5B (monosomic 5B) can produce F₁ hybrids deficient for Ph1, facilitating recombination in those specific plants.
- Substitutions: Replacing chromosome 5B with a homoeologue, such as rye chromosome 5R, is another method used to achieve Ph1 deficiency.
4. Enhancement through Promoter Genes
Recent research has identified additional genes that can be used in conjunction with Ph1 manipulation to boost success rates:
- Hpp-5Mg: Found on chromosome 5MgS of Ae. geniculata, this gene acts as a homoeologous pairing promoter. When combined with the ph1b mutation, it can increase the frequency of recombinant chromosomes many-fold and even promote recombination in the usually recalcitrant proximal (centromeric) regions of chromosomes.
Strategic Outcomes of Manipulation
Manipulating the Ph1 locus allows for “chromosome engineering,” which is the high-precision transfer of small alien segments rather than entire arms or chromosomes. This is critical for minimizing linkage drag, where undesirable wild traits (like poor yield or quality) remain linked to the beneficial target gene.
Once the desired transfer is achieved, breeders must eliminate the ph1b mutant chromosome or the suppressor genes by backcrossing the lines with wild-type wheat. This restores the functional Ph1 system, ensuring that the newly modified genome is cytologically stable for use in commercial agricultural production.
14. What are the trade-offs of using rye in wheat breeding?
The use of rye (Secale cereale) in wheat breeding presents a complex balance between significant agronomic gains and notable industrial drawbacks. While rye introgressions are the most widely utilized alien genetic resources for wheat improvement, they require careful management to mitigate negative impacts on flour performance and genetic stability.
Primary Benefits of Rye Introgressions
Rye chromosomes, particularly segments from 1R, 2R, and 3R, offer a robust suite of protective and yield-enhancing traits:
- Disease Resistance: The most famous introgression, the 1RS chromosome arm, carries a “supergene” block providing resistance to multiple fungal diseases, including stem rust (Sr31), stripe rust (Yr9), leaf rust (Lr26), and powdery mildew (Pm8). Novel genes like Sr59 from chromosome 2R remain effective against highly virulent stem rust races like Ug99.
- Abiotic Stress Tolerance: Rye chromatin significantly improves wheat’s resilience to drought. This is largely due to more robust and deeper root systems associated with the 1RS arm, which allow for better water accessibility in deep soil layers. Chromosome 3R also contributes tolerance to aluminum toxicity.
- Yield and Vigor: Introgressions like 1BL.1RS have contributed to a great increase in global yield potential and aerial biomass.
- Nutritional and Sustainability Traits: Rye introgressions can enhance levels of Zinc (Zn) and Iron (Fe) in grain and provide allelopathic potential, which helps the crop naturally suppress weed growth.
Critical Trade-offs and Drawbacks
Despite these advantages, breeders face several challenges when using rye genetic material:
- Negative Impact on Baking Quality: This is the most significant trade-off. The widely used 1RS translocation often results in weak and sticky dough. This is caused by a “double effect”: the deletion of essential wheat glutenin proteins (Glu-B3/Gli-B1) and the introduction of rye secalins (Sec-1 locus) that interfere with dough handling and polymerization.
- Note: Recent research suggests introgressions from chromosome 3R may offer a more favorable trade-off by contributing to baking quality stability under drought without causing the stickiness associated with 1RS.
- Linkage Drag: Because rye genes are often transferred as large chromosomal segments, beneficial alleles are frequently linked to deleterious wild traits that can negatively affect yield or quality if not carefully pruned through advanced chromosome engineering.
- Meiotic and Genomic Instability: Introgressed rye chromosomes often exhibit reduced meiotic pairing compared to the host wheat genome. This instability can lead to the gradual elimination of the rye material over successive generations unless selection pressure is maintained. This is often due to a failure of rye telomeres to properly attach to the nuclear envelope and cluster during the onset of meiosis.
- Vulnerability to Evolving Pathogens: Although rye provides broad resistance, some heavily deployed genes, such as Sr31, have eventually been overcome by new virulent races of pathogens, leading to “boom and bust” cycles in monocultures.
In summary, while rye is an indispensable tool for securing wheat production under climate change and disease pressure, the primary trade-off involves a compromise in end-use processing quality and the requirement for sophisticated breeding techniques to maintain genomic stability.
15. Which Aegilops species best improve baking quality?
Several Aegilops species, commonly known as goatgrasses, are recognized as superior genetic donors for improving the baking and bread-making quality of wheat. These wild relatives contribute novel alleles for seed storage proteins—specifically high-molecular-weight glutenin subunits (HMW-GS) and low-molecular-weight glutenin subunits (LMW-GS)—and grain texture genes that were often lost during wheat domestication.
Top Aegilops Species for Baking Quality
- Aegilops geniculata (Mg and Ug genomes): This species is one of the most promising donors. Chromosome 1Mg significantly increases grain hardness, protein content, gluten content, and Zeleny sedimentation value. Additionally, chromosome 1Ug introgressions enhance the rheological properties of dough by improving the microstructure of the gluten network. It also has a positive effect on seed volume-weight.
- Aegilops longissima (Sl genome): Chromosome 1Sl is renowned for carrying genes that enhance bread-making quality. Wheat lines with 1Sl substitutions have shown improved farinograph properties and higher bread-making scores compared to standard wheat cultivars.
- Aegilops umbellulata (U genome): This relative provides LMW-GS from the 1U genome that confer superior dough rheology and overall bread-making quality. Specific LMW-GS substitution lines (Glu U3a and Glu U3b) have been shown to increase mixing properties and loaf volume.
- Aegilops searsii (Ss genome): The addition of the 1Ss chromosome has been reported to increase both protein content and gluten strength. It provides a novel pair of HMW-GS that improves the end-product quality of hexaploid wheat.
- Aegilops biuncialis (Ub and Mb genomes): The 1Ub addition line carries HMW-GS directly associated with high baking quality. Furthermore, 1Mb(1B) substitution lines have also demonstrated favorable quality-associated glutenin subunits.
- Aegilops caudata (C genome): Specific substitution lines (1C(1A)) carrying novel HMW-GS (1Dx2s and 1Dx2f) have shown superior gluten strength compared to the bread wheat landrace Chinese Spring. It also improves dough development and stability time.
- Aegilops sharonensis (Ssh genome): This species contributes HMW-GS with unusually large molecular weights, which have been shown to improve bread-making quality in wheat-Ae. sharonensis introgression lines.
Specialized Quality Traits
- Extra-Soft Texture: Aegilops triuncialis is a valuable donor for the extra-soft grain texture trait. The introgression of chromosome 5U (specifically 5U-5A substitutions) introduces functional puroindoline genes (Pina and Pinb) that reduce grain hardness, making the flour ideal for biscuits and cookies.
- Grain Morphology: Chromosome 1M from Aegilops comosa has been reported to have a positive effect on seed morphology alongside its contributions to bread-making quality.
Summary of Quality Enhancements
| Aegilops Species | Target Chromosome | Primary Phenotypic Effect |
|---|---|---|
| Ae. geniculata | 1Mg, 1Ug | Increased hardness, gluten strength, and dough rheology. |
| Ae. longissima | 1Sl | Enhanced bread-making scores and farinograph stability. |
| Ae. umbellulata | 1U (LMW-GS) | Improved dough development time and loaf volume. |
| Ae. searsii | 1Ss | Improved protein quality and gluten strength. |
| Ae. biuncialis | 1Ub, 1Mb | Favorable baking-quality associated HMW-GS. |
| Ae. triuncialis | 5U | Increased grain softness for biscuit-making. |
| Ae. caudata | 1C | Superior dough development and stability. |
16. What is the allelopathic potential of rye in organic wheat?
Rye (Secale cereale) is considered an excellent source of allelopathic potential, which refers to a plant’s ability to naturally suppress weed growth through the release of biochemicals. This trait is particularly valuable for organic wheat farming, where herbicide use is restricted and managing weed competition is a significant challenge.
Key aspects of rye’s allelopathic potential in wheat breeding include:
- Weed Suppression: Wheat-rye introgression lines have demonstrated the ability to suppress various weeds, including Sinapis alba (mustard), while standard bread wheat genotypes often show no such allelopathic activity.
- Chromosomal Origins: The highest allelopathic potential has been identified in lines carrying rye chromosomes 1R and 2R. Additionally, lines with multiple rye chromosomes, such as 1R + 6R and the complex combination 1R + 4R + 6R + 7R, also exhibit high activity.
- Early Vigor: Research indicates that wheat lines with a 1R substitution often show early growth vigor. This vigor is believed to be positively linked to the exudation of allelochemicals from the roots, enhancing the plant’s competitive edge against weeds during early development.
- Quantitative Inheritance: Allelopathic activity is a complex, quantitatively inherited trait. While it is more difficult to breed than traits governed by a single gene, successful breeding for improved allelopathy has been achieved in other crops like rice and spring wheat.
By utilizing these wheat-rye introgression lines, breeders aim to develop allelopathic wheat cultivars that offer a sustainable solution for weed management in organic and low-input agricultural systems.
17. Which Aegilops chromosomes increase total seed protein content?
Several species within the Aegilops genus (goatgrasses) possess chromosomes that, when introgressed into bread wheat, significantly increase total seed protein content and improve related nutritional qualities. These improvements are primarily driven by novel alleles for seed storage proteins and genes regulating nutrient remobilization.
Aegilops geniculata (Mg and Ug genomes)
This species is one of the most productive donors for protein enhancement.
- Chromosomes 1Mg and 1Ug: Introgressions of 1Mg have a major positive effect on total seed protein concentration, gluten content, and grain hardness. 1Ug also contributes to protein and improves dough rheology.
- Group 2, 4, 5, and 7 Chromosomes: Introgressions of 2Mg, 7Mg, 2Ug, 4Ug, 5Ug, and 7Ug have all been reported to increase total seed protein content in bread wheat.
- 5Mg: Specifically, 5Mg contributes to increased grain protein while also positively affecting dietary fiber and beta-glucan content.
Aegilops biuncialis (Mb and Ub genomes)
Aegilops biuncialis introgressions are effective for boosting the nutritional value of wheat flour.
- Group 2 and 3 Mb Chromosomes: Chromosome 2Mb and 3Mb are known to enhance grain protein content in wheat-Aegilops addition lines.
- Group 5 and 7 Mb/Ub Chromosomes: The addition of Mb and Ub chromosomes from groups 5 and 7 to wheat results in a significant increase in both grain protein and health-promoting dietary fibers.
Aegilops kotschyi and Aegilops peregrina (S and U genomes)
These species are valued for providing a combined boost to protein and essential minerals.
- Ae. kotschyi Group 2 and 7: High grain protein concentration in these introgression lines is strongly correlated with the presence of 2Sk, 7Uk, or 7Sk chromosomes.
- Ae. kotschyi Group 1: The 1Uk and 1Sk chromosomes have demonstrated the greatest effect on increasing micronutrients and protein, with some derivatives showing a 33% increase in protein content.
- Ae. peregrina Groups 2, 4, and 7: Introgressions from chromosomes 2Sv, 2Up, 4Sv, 4Up, 7Sv, and 7Up are associated with significantly higher protein, Zinc (Zn), and Iron (Fe) levels.
Other Aegilops Donors
- Aegilops searsii (Ss genome): The addition of chromosome 1Ss increases both total protein content and gluten strength.
- Aegilops longissima (Sl genome): Chromosomes 1Sl and 2Sl are recognized for enhancing protein properties and grain micronutrient concentration.
- Aegilops tauschii (D genome): Although common wheat already contains the D genome, novel quantitative trait loci (QTLs) for increased grain protein content (GPC) have been successfully transferred from wild Ae. tauschii accessions into synthetic wheat lines.
Summary of Protein-Enhancing Aegilops Chromosomes
| Species | Chromosomes Linked to Increased Protein | Additional Benefits |
|---|---|---|
| Ae. geniculata | 1Mg, 2Mg, 7Mg, 1Ug, 2Ug, 4Ug, 5Ug, 7Ug | Grain hardness, gluten strength, high Fe/Zn. |
| Ae. biuncialis | 2Mb, 3Mb, 5Mb, 7Mb, 7Ub | Dietary fiber and beta-glucan content. |
| Ae. kotschyi | 1Sk, 1Uk, 2Sk, 7Sk, 7Uk | Massive increases in Iron and Zinc content. |
| Ae. peregrina | 2Sv, 2Up, 4Sv, 4Up, 7Sv, 7Up | High salt and drought tolerance. |
| Ae. searsii | 1Ss | Enhanced baking quality. |
| Ae. longissima | 1Sl, 2Sl | Improved end-use quality scores. |
16. Do these Aegilops introgressions affect health-promoting dietary fibers?
Yes, Aegilops introgressions have a significant impact on the levels of health-promoting dietary fibers in wheat, offering a way to restore genetic diversity lost during domestication. These fibers, including polysaccharides like arabinoxylan and beta-glucan, are essential for human health, providing protection against cardiovascular diseases, type 2 diabetes, and colorectal cancer.
Key Aegilops Species and Chromosomes
Research has identified specific species and chromosomal segments that positively influence fiber content:
- Aegilops biuncialis: The addition of group 5 and group 7 chromosomes from this species to wheat results in a significant increase in both grain protein and dietary fiber content, improving the overall nutritional value of the flour.
- Aegilops geniculata: Introgressions from chromosomes 5Ug and 5Mg have been shown to have a positive effect on both the dietary fiber composition and the beta-glucan content of hexaploid wheat.
- Other Potential Donors: Broad screening of Aegilops species (including genomes U, S, and D) has highlighted chromosomes 2S, 2U, 7S, and 7U as additional sources for enhancing dietary fiber levels.
Nutritional and Functional Implications
- Functional Foods: Because these introgressions (particularly the 5Mb(5D) substitution from Ae. biuncialis) boost the concentration of health-related components, they are considered valuable genetic resources for the production of functional foods.
- Bioactive Compounds: Beyond standard fibers, some Aegilops germplasms also exhibit elevated levels of other health-promoting bioactive compounds, such as phenolic compounds and various vitamins.
- Bread-making Trade-offs: While increasing dietary fiber is beneficial for nutrition, it can alter processing characteristics; for instance, high beta-glucan content is known to increase dough viscosity and water absorption while potentially reducing loaf volume.
In summary, the significant variability for dietary fibers found in Aegilops species represents a rich reservoir that breeders are actively exploiting to develop elite wheat cultivars with enhanced nutritional profiles.
17. Which other wild relatives offer high iron and zinc?
Beyond the species already discussed, several other wild relatives of wheat are recognized as high-value reservoirs for Iron (Fe) and Zinc (Zn), often containing concentrations two to three times higher than cultivated varieties.
Aegilops (Goatgrass) Species
The Aegilops genus is a primary target for biofortification due to its vast natural variation in micronutrients.
- Aegilops kotschyi: This species is one of the most effective donors, with some accessions exhibiting three to fourfold higher levels of iron and zinc than modern wheat. Introgressions of its 1Uk, 1Sk, 2Sk, 7Uk, and 7Sk chromosomes have been shown to significantly boost grain mineral density.
- Aegilops peregrina: Chromosomes from groups 4 and 7 of this species are associated with significantly higher Iron and Zinc levels. It has been used to develop addition lines that double the micronutrient content of recipient wheat varieties.
- Aegilops longissima: Recognized for its potential to improve grain nutritional quality, particularly via chromosomes 1Sl, 2Sl, and 7Sl.
- Aegilops tauschii: As the donor of wheat’s D genome, it is a rich reservoir for Fe and Zn uptake genes.
- Other Aegilops donors: Ae. caudata (1C), Ae. searsii (1Ss, 2Ss), and Ae. umbellulata (2U, 6U) have all been identified as viable sources for increasing these essential minerals.
Wild Triticum Relatives
- Wild Emmer (Triticum dicoccoides): This is perhaps the most famous donor due to the GPC-B1 (NAM-B1) gene on chromosome 6BS. This locus not only increases grain protein but also significantly enhances the remobilization of Zinc, Iron, and Manganese to the grain.
- Einkorn Species (T. monococcum and T. boeoticum): These wild diploid wheats have been reported to carry significant variation for high grain Iron and Zinc content.
Perennial Grasses
- Leymus Species: Crosses with Leymus racemosus and Leymus mollis have produced introgression lines with high levels of Zinc, often combined with low Cadmium concentrations.
- Thinopyrum junceiforme: This species has been identified as a source for increased Zinc content in wheat-alien hybrids.
Rye (Secale cereale)
While primarily used for disease resistance, rye is also a valuable source for micronutrients. Wheat lines with introgressions of rye chromosomes 1R, 2R, 3R, and 5R have demonstrated elevated Iron and Zinc concentrations, with Zinc levels reaching as high as 89.3 mg/kg in some experimental lines.
18. What causes the ‘double effect’ on dough quality?
The “double effect” on dough quality refers to the simultaneous genetic loss and alien protein introduction that occurs during the widespread 1BL.1RS wheat-rye translocation. This exchange significantly compromises the baking and processing characteristics of wheat flour through two distinct mechanisms:
- Deletion of Critical Wheat Proteins: When the 1RS rye chromosome arm replaces the 1BS wheat arm, the Glu-B3 and Gli-B1 loci are deleted. These loci are essential because they encode low-molecular-weight (LMW) glutenin subunits and gliadins that provide structural integrity and elasticity to the gluten network. Their absence results in reduced dough strength, often referred to as “weak dough“.
- Introduction of Rye Secalins: The introgressed 1RS arm brings with it the Sec-1 locus, which encodes rye storage proteins known as γ-secalin and ω-secalin. These high-molecular-weight proteins interfere with the standard polymerization of wheat gluten. Specifically, the presence of these secalins is the primary cause of increased dough stickiness, making the dough difficult to handle during industrial processing.
Summary of the Impact
| Component | Genetic Change | Phenotypic Result |
| Wheat 1BS Arm | Deletion of Glu-B3/Gli-B1 | Weak dough due to loss of glutenin/gliadin. |
| Rye 1RS Arm | Addition of Sec-1 locus | Sticky dough caused by γ– and ω-secalins. |
While this “double effect” traditionally limits the use of 1RS in premium baking wheats, researchers have found that dough functionality can be improved if gluten protein genes are restored within these translocation lines. Additionally, introgressions from other rye chromosomes, such as 3R, have been found to bolster protein concentration without causing the stickiness associated with the 1RS arm.
19. How is the Ph1 locus manipulation reversed after breeding?
After the desired alien genetic material has been successfully introgressed into the wheat genome, the manipulation of the Ph1 locus must be reversed to stabilize the genome and ensure regular meiotic behavior for commercial use.
The reversal is achieved through the following steps:
1. Backcrossing with Wild-Type Wheat
The primary method for reversing the manipulation is to backcross the recombinant lines with wild-type wheat (parents that carry the functional, dominant Ph1 gene). This process introduces a functional chromosome 5B to replace the ph1b mutant chromosome or to remove external suppressor genes like those derived from Aegilops speltoides.
2. Marker-Assisted Selection (MAS)
Because it is impossible to distinguish between plants that are homozygous for the functional Ph1 gene and those that are hemizygous (carrying one functional and one mutant copy) through appearance alone, breeders use molecular markers. Specific DNA markers, such as PSR128 and PSR574, are used to:
- Confirm the reintroduction of the functional Ph1 gene.
- Identify and eliminate the ph1b deletion segment from the selected offspring.
- Ensure the recombinant line is homozygous for the functional Ph1 locus, which usually requires additional generations of selection and testing.
3. Genome Stabilization
Restoring the functional Ph1 locus is critical because it acts as a genetic “lock.” Once restored, it prevents the newly introgressed alien segments from undergoing further homoeologous recombination. This ensures that the alien gene behaves as a single, stable locus that will be inherited predictably in future generations.
Without this reversal, the wheat genome would remain cytologically unstable, leading to irregular chromosome pairing, reduced fertility, and the potential loss of the beneficial alien trait during subsequent reproduction.
20. How do irradiation techniques help minimize linkage drag?
Irradiation techniques, such as the use of X-rays or gamma-rays, help minimize linkage drag by inducing random chromosomal damage and breaks that can separate a target gene from nearby undesirable alien chromatin. This is particularly critical when dealing with wild relatives from the secondary or tertiary gene pools, whose chromosomes may not naturally pair or recombine with the wheat genome during meiosis.
Mechanism for Reducing Linkage Drag
- Inducing Random Breaks: Ionizing radiation causes random physical breaks in both the wheat and alien donor chromosomes. When these fragments fuse, they form translocations. Because the breaks are random, they can occur between a target gene and a tightly linked deleterious gene, effectively “breaking” the linkage that conventional recombination cannot.
- Transfer of Small Segments: By facilitating these random breaks, irradiation allows for the translocation of very small alien chromosome fragments into the wheat genome. These small segments are less likely to contain undesirable wild traits—such as poor yield or baking quality—that typically accompany the introgression of entire chromosome arms.
- Bypassing Pairing Barriers: In cases where meiotic homoeologous pairing is blocked (such as by the Ph1 locus) or simply impossible due to lack of genomic similarity, irradiation serves as a physical alternative to induce the necessary genetic rearrangements.
Methodologies and Applications
- Pollen Irradiation: One strategy involves irradiating immature pollen or dehiscing anthers. This irradiated pollen is then used to fertilize emasculated wheat spikes, resulting in hybrid derivatives that carry diverse chromosomal rearrangements.
- Seed Irradiation: Researchers have also successfully used seed irradiation to transfer useful variability, such as high grain Iron and Zinc content from Aegilops kotschyi into adapted wheat varieties.
- Historical Precision: A pioneering example of this technique was E.R. Sears’ transfer of the Lr9 leaf rust resistance gene from Ae. umbellulata. By using X-rays, Sears produced over 17 different translocations; from these, he selected a specific, cytologically undetectable “cryptic” translocation that carried the resistance with minimal negative side effects.
While highly effective for breaking linkages, irradiation is a random process that often requires screening very large populations to find a “breeder-friendly” translocation that is both stable and has the smallest possible alien segment.
21. What breeding techniques best mitigate linkage drag from rye?
To mitigate linkage drag from rye—where beneficial traits like disease resistance are genetically tied to undesirable genes like those for sticky dough—breeders primarily use techniques that facilitate the transfer of small, targeted alien segments rather than entire chromosome arms.
The most effective techniques for this purpose include:
1. Manipulation of the Ph1 Locus (Induced Homoeologous Recombination)
The ph1b mutant is the most widely utilized tool for breaking the genetic barriers between wheat and rye.
- Mechanism: In its wild-type state, the Ph1 gene restricts pairing to identical partners. Disabling it allows rye chromosomes to synapsis and recombine with their wheat homoeologues.
- Outcome: This facilitates chromosome engineering, where breeders can “prune” a large rye translocation (like the 1RS arm) down to a much smaller segment that carries the target gene but excludes deleterious genes like the Sec-1 locus (responsible for sticky dough secalins).
2. Ionizing Irradiation
Irradiation with X-rays or gamma-rays is a physical method used to induce random chromosomal damage.
- Breaking Linkage: Because the breaks caused by radiation are random, they can occur between a target gene and a tightly linked undesirable trait that conventional recombination cannot separate.
- Precise Transfers: This technique has been historically successful in creating “cryptic” translocations that are cytologically undetectable but carry essential resistance with minimal negative side effects.
3. DNA Marker-Assisted Chromosome Engineering (MACE)
Advanced genomic tools are now used to guide and verify the reduction of alien chromatin.
- Marker Monitoring: High-throughput genotyping—such as SNP arrays, GBS (Genotyping-by-Sequencing), and KASP markers—allows breeders to monitor the presence of rye chromatin in large populations.
- Selection for Small Segments: Markers flanking a target gene enable breeders to select specific recombinants that have lost the maximum amount of “excess” rye DNA while retaining the beneficial allele.
4. Secondary and Tertiary Recombination
Linkage drag can be further minimized by inducing multiple rounds of recombination.
- Pruning Process: A primary wheat–rye recombinant is crossed back to wheat to induce secondary recombination.
- Increased Homology: Because the primary recombinant segment already shares more similarity with the wheat chromosome than the original rye chromosome, it facilitates more frequent and precise crossovers in subsequent generations.
5. Precision Genome Editing (CRISPR-Cas9)
While still an emerging field in wheat breeding, CRISPR-Cas9 offers the potential to eliminate linkage drag entirely.
- Allele Replacement: Rather than introgressing a large piece of alien chromatin, CRISPR can be used to precisely mirror rye alleles within the wheat genome or replace endogenous wheat genes with their rye-derived counterparts.
- Deactivating Drag: It can also be used to selectively deactivate undesirable genes (like the Sec-1 loci) within an existing translocation line to improve its processing quality without losing resistance.
22. What chromosome engineering steps followed the Ph1 manipulation?
Following the initial manipulation of the Ph1 locus to induce homoeologous pairing, breeders must execute several critical chromosome engineering steps to recover, stabilize, and refine the alien introgression. These steps move the genetic material from unstable hybrid states into “breeder-friendly” lines suitable for commercial use.
1. Recovery and Identification of Recombinants
Once recombination has been induced (e.g., in a hybrid homozygous for ph1b), the next step is recovering those specific events in the progeny.
- The Top-Cross Strategy: In this method, synthetic hexaploids are top-crossed with hexaploid wheat to produce TC1 plants, which are double monosomic for the alien and D-genomes. These are then either self-pollinated or top-crossed again to generate a TC2F1 population.
- Cytogenetic Screening: Advanced techniques like GISH (Genomic In Situ Hybridization) and FISH (Fluorescence In Situ Hybridization) are used to detect the presence of alien segments and identify the specific chromosomes involved in the rearrangement.
- Marker-Assisted Selection (MAS): High-throughput tools, such as 90K SNP arrays and PCR-based markers, are employed to screen large populations for recombinant gametes that carry the target gene without the need for laborious microscopy at every stage.
2. Selection of Disomic Lines
Most primary wheat–alien translocations initially occur in monosomic form.
- Stabilizing Inheritance: Breeders must select for disomic translocation lines (carrying two copies of the recombinant chromosome) to ensure stable inheritance across future generations.
- Agronomic Evaluation: Disomic lines are essential for accurately assessing the alien chromatin’s impact on wheat yield and quality, as monosomic lines are often cytologically unstable.
3. Elimination of the ph1b Mutation
A critical subsequent step is the restoration of the functional Ph1 gene.
- Backcrossing: The recombinant lines are backcrossed with wild-type wheat (carrying the functional Ph1 gene) to eliminate the ph1b mutant chromosome.
- Genome “Locking”: Reintroducing the functional Ph1 locus acts as a genetic lock that prevents the newly introgressed alien segments from undergoing further homoeologous recombination, thereby stabilizing the genome for elite breeding programs.
4. Linkage Drag Reduction (Pruning)
If the initially introgressed alien segment is large, it may still carry undesirable traits (linkage drag).
- Secondary and Tertiary Recombination: Breeders may induce additional rounds of recombination to “prune” the alien segment down to the smallest possible size that still retains the target gene.
- Improved Homology: Because the primary recombinant segment already shares more similarity with the wheat chromosome than the original alien chromosome, it facilitates more frequent and precise crossovers in these subsequent generations.
- Directed Engineering: This process aims to produce “cryptic” translocations—segments so small they are sometimes cytologically undetectable but carry the desired trait with minimal negative side effects.
23. Can CRISPR-Cas9 eliminate linkage drag from rye secalins?
Yes, CRISPR-Cas9 is considered a promising alternative and potentially the “method of choice” for eliminating the linkage drag associated with rye secalins.
The primary issue with the widely used 1RS translocation is that beneficial disease resistance genes (like Sr31 and Yr9) are genetically linked to the Sec-1 locus, which encodes γ-secalin and ω-secalin. These proteins are responsible for the weak and sticky dough performance that reduces bread-making quality.
According to the sources, CRISPR-Cas9 can address this drag through several precision strategies:
- Deactivating Deleterious Genes: Linkage drag can be mitigated by selectively deactivating undesired genes, such as the Sec-1 loci, within an existing translocation line while leaving the beneficial resistance genes intact.
- Allele Replacement and Mirroring: Rather than transferring a large segment of alien chromatin that inevitably contains “drag,” CRISPR can be used to replace endogenous wheat alleles with specific beneficial alien alleles or to introduce “mirror” mutations that replicate alien traits within the wheat genome.
- Multiplexing for Complex Traits: CRISPR systems can target multiple genomic loci simultaneously, allowing for the rapid introgression of complex trait combinations without the need for the large, deleterious segments used in traditional homoeologous recombination.
While traditional methods like ph1b-induced recombination or irradiation are used to “prune” alien segments, they are often laborious and may only localize recombination to specific regions. Precision genome editing provides a more targeted approach to severing the link between high yield/resistance and poor processing quality.
References
Abrouk, M., Wang, Y., Cavalet-Giorsa, E., Troukhan, M., Kravchuk, M., & Krattinger, S. G. (2023). Chromosome-scale assembly of the wild wheat relative Aegilops umbellulata. Scientific Data, 10(1). https://doi.org/10.1038/s41597-023-02658-2
Ali, N., Heslop-Harrison, J., Ahmad, H., Graybosch, R. A., Hein, G. L., & Schwarzacher, T. (2016). Introgression of chromosome segments from multiple alien species in wheat breeding lines with wheat streak mosaic virus resistance. Heredity, 117(2), 114–123. https://doi.org/10.1038/hdy.2016.36
Ara, A. (2018). Alien Gene Introgression in Crop Improvement – New Insights. International Journal of Pure & Applied Bioscience, 6(6), 890–904. https://doi.org/10.18782/2320-7051.7195
Badaeva, E. D., Davoyan, R. O., Tereshchenko, N. A., Lyalina, E. V., Zoshchuk, S. A., & Goncharov, N. P. (2024). Cytogenetic features of intergeneric amphydiploids and genome-substituted forms of wheat. Vavilovskii Zhurnal Genetiki i Selektsii, 28(7), 716–730. https://doi.org/10.18699/vjgb-24-80
Badaeva, E. D., González Franco, M. J., Razumova, O., Tereshchenko, N. A., & Divashuk, M. (2025). Perspectives on the utilization of Aegilops species containing the U genome in wheat breeding: a review. In Frontiers in Plant Science (Vol. 16). Frontiers Media SA. https://doi.org/10.3389/fpls.2025.1661257
Boehm, J., & Cai, X. (2024). Enrichment and Diversification of the Wheat Genome via Alien Introgression. In Plants (Vol. 13, Number 3). Multidisciplinary Digital Publishing Institute (MDPI). https://doi.org/10.3390/plants13030339
Brar, D. S., & Khush, G. S. (1997). Alien introgression in rice. In Plant Molecular Biology (Vol. 35).
Dong, Z., Ma, C., Tian, X., Zhu, C., Wang, G., Lv, Y., Friebe, B., Li, H., & Liu, W. (2020). Genome-wide impacts of alien chromatin introgression on wheat gene transcriptions. Scientific Reports, 10(1). https://doi.org/10.1038/s41598-020-61888-1
Farkas, A., Gaál, E., Ivanizs, L., Blavet, N., Said, M., Holušová, K., Szőke-Pázsi, K., Spitkó, T., Mikó, P., Türkösi, E., Kruppa, K., Kovács, P., Darkó, É., Szakács, É., Bartoš, J., Doležel, J., & Molnár, I. (2023). Chromosome genomics facilitates the marker development and selection of wheat-Aegilops biuncialis addition, substitution and translocation lines. Scientific Reports, 13(1). https://doi.org/10.1038/s41598-023-47845-8
Gavrilenko, T., Pendinen, G., Antonova, O., Makarova, T., & Thieme, R. (2023). Homoeologous Chromosome Pairing and Alien Introgression in Backcrossing Progenies Derived from Hybrids Solanum tuberosum (+) Mexican 2x (1 EBN) B-Genome Potato Species. Agronomy, 13(7). https://doi.org/10.3390/agronomy13071809
Gill, B. S., Friebe, B. R., & White, F. F. (2011). Alien introgressions represent a rich source of genes for crop improvement. In Proceedings of the National Academy of Sciences of the United States of America (Vol. 108, Number 19, pp. 7657–7658). https://doi.org/10.1073/pnas.1104845108
Johansson, E., Henriksson, T., Prieto-Linde, M. L., Andersson, S., Ashraf, R., & Rahmatov, M. (2020). Diverse Wheat-Alien Introgression Lines as a Basis for Durable Resistance and Quality Characteristics in Bread Wheat. Frontiers in Plant Science, 11. https://doi.org/10.3389/fpls.2020.01067
Johansson, E., Lan, Y., Olalekan, O., Kuktaite, R., Chawade, A., & Rahmatov, M. (2024). Alien introgression to wheat for food security: functional and nutritional quality for novel products under climate change. In Frontiers in Nutrition (Vol. 11). Frontiers Media SA. https://doi.org/10.3389/fnut.2024.1393357
Kovács, P., Farkas, A., Türkösi, E., Kruppa, K., Szakács, É., Szőke-Pázsi, K., Hidvégi, N., Mikó, P., Gulyás, A., Darkó, É., Said, M., Ivanizs, L., Gaál, E., & Molnár, I. (2026). A genetransfer strategy based on durum wheat–Aegilops comosa amphiploid top-crossed with hexaploid wheat results in elimination of D chromosomes led to the production of homoeologous M(D) substitutions and translocations. BMC Plant Biology. https://doi.org/10.1186/s12870-025-07950-4
Namazova, L. (2024). CYTOGENETIC AND AGRO-MORPHOLOGICAL STUDY OF SUBSTITION LINES BETWEEN BREAD WHEAT AND AEGILOPS UMBELLULATA. Research in: Agricultural & Veterinary Sciences, 8(3), 177–184. https://doi.org/10.62476/ravs83177
Perničková, K., Koláčková, V., Lukaszewski, A. J., Fan, C., Vrána, J., Duchoslav, M., Jenkins, G., Phillips, D., Šamajová, O., Sedlářová, M., Šamaj, J., Doležel, J., & Kopecký, D. (2019). Instability of alien chromosome introgressions in wheat associated with improper positioning in the nucleus. International Journal of Molecular Sciences, 20(6). https://doi.org/10.3390/ijms20061448
Rey, E., Molnár, I., & Doležel, J. (2015). Genomics of wild relatives and alien introgressions. In Alien Introgression in Wheat: Cytogenetics, Molecular Biology, and Genomics (pp. 347–381). Springer International Publishing. https://doi.org/10.1007/978-3-319-23494-6_13
Saini, P., Kaur, H., Tyagi, V., Yadav, A. N., Saini, P., Sharma, V., Singh, C., Dhaliwal, H. S., & Sheikh, I. (2023). Genetic enhancement of nutritional and end-use quality in bread wheat through alien introgressions from wild relatives. In Cereal Research Communications (Vol. 51, Number 2, pp. 295–314). Akademiai Kiado ZRt. https://doi.org/10.1007/s42976-022-00309-7
Singh, J. (n.d.). DEVELOPMENT OF GENOMIC RESOURCES AND EXPLORATION OF DISEASE RESISTANCE IN AEGILOPS UMBELLULATA TO IMPROVE THE CULTIVATED WHEAT.
Sobhakumari, V. P., Mohanraj, K., Nair, N. V., Mahadevaswamy, H. K., & Ram, B. (2020). Cytogenetic and molecular approaches to detect alien chromosome introgression and its impact in three successive generations of erianthus procerus × saccharum. Cytologia, 85(4), 341–346. https://doi.org/10.1508/CYTOLOGIA.85.341
Yunusbaev, U., Campos, G. R., Sathishraj, R., Liakh, E., Raupp, J. W., Zimin, A. V., Akhunova, A., Koo, D. H., & Akhunov, E. (2025). The Evolution and Origin of Allotetraploid Aegilops geniculata Revealed by the Homoeolog-Resolved Genome Assembly. Plant Biotechnology Journal. https://doi.org/10.1111/pbi.70456
Zhang, S., Zhang, Y., Hu, T., Li, L., Wang, Z., Qiao, L., Chang, L., Li, X., Chang, Z., Zhang, P., & Zhang, X. (2025). Impact of Alien Chromosome Introgression from Thinopyrum ponticum on Wheat Grain Traits. Plants, 14(19). https://doi.org/10.3390/plants14193072
Zhang, W., Cao, Y., Zhang, M., Zhu, X., Ren, S., Long, Y., Gyawali, Y., Chao, S., Xu, S., & Cai, X. (2017). Meiotic homoeologous recombination-based alien gene introgression in the genomics era of wheat. Crop Science, 57(3), 1189–1198. https://doi.org/10.2135/cropsci2016.09.0819
Related posts:
 The Evolutionary Genomics of Crop Plants: From Ancestral Polyploidy to Modern Pangenomes
The Evolutionary Genomics of Crop Plants: From Ancestral Polyploidy to Modern Pangenomes
 The Secret of the Seed: An Aspiring Learner’s Guide to Hybrid Vigor and Plant Development
The Secret of the Seed: An Aspiring Learner’s Guide to Hybrid Vigor and Plant Development
 Comprehensive Guide to Cytoplasmic Male Sterility: Molecular Mechanisms, ncRNA Regulation, and Advanced Genomic Editing for Crop Improvement
Comprehensive Guide to Cytoplasmic Male Sterility: Molecular Mechanisms, ncRNA Regulation, and Advanced Genomic Editing for Crop Improvement
 The Evolution of Plant Mutagenesis: Advanced Methodologies for Global Crop Improvement and Food Security
The Evolution of Plant Mutagenesis: Advanced Methodologies for Global Crop Improvement and Food Security
 The Architecture of Yield: A Comprehensive Guide to Plant Chromosome Engineering and CRISPR-Mediated Genomic Innovation
The Architecture of Yield: A Comprehensive Guide to Plant Chromosome Engineering and CRISPR-Mediated Genomic Innovation