1. Introduction to Next-Generation Vector Control
The eradication of malaria and the management of invasive species represent two of the most critical imperatives in modern global health and conservation biology. Traditional vector control strategies, while foundational, often suffer from diminishing returns due to insecticide resistance and logistical scaling issues. Gene drive technology offers a strategic paradigm shift, providing a mechanism to either suppress vector populations or modify them to be refractory to disease transmission within a deterministic prediction of only a few years.

At their core, gene drives are “selfish genetic elements” that bypass standard Mendelian inheritance. While a standard allele possesses a 50% probability of transmission, gene drives utilize super-Mendelian mechanisms to ensure they are inherited at frequencies approaching 100%. The “CRISPR revolution” has radically simplified the engineering of these elements. Unlike older technologies like zinc-finger nucleases or TALENs, which required laborious protein engineering for every new target, the CRISPR-Cas9 system uses a programmable guide RNA (gRNA) to direct the Cas9 endonuclease to a specific protospacer sequence. This flexibility allows for the rapid development of functional laboratory systems, yet the operational success of these drives is dictated by the precise molecular mechanisms of repair and the subsequent emergence of genetic resistance.

1.1 The Genetic Baseline: Mendelian vs. Gene Drive Inheritance
In the traditional classroom, we teach that inheritance is a 50/50 coin flip. This is Mendelian inheritance: each offspring has a 50% chance of receiving a specific allele from a parent. However, in the world of genomic engineering, we have unlocked “Super-Mendelian” inheritance.
A CRISPR gene drive is a genetic system that “cheats” these rules. By actively copying itself onto the opposing chromosome, a drive can achieve transmission rates of 91.4% to 99.6%, as observed in landmark studies on the Anopheles gambiae mosquito.
“Aha” Moment: Think of a gene drive like a race where one runner has a special power: mid-race, they can touch any other runner and instantly turn them into an identical copy of themselves. Soon, every person crossing the finish line is that same runner.
| Inheritance Patterns | Standard Mendelian Genetics | CRISPR Gene Drive |
|---|---|---|
| Primary Mechanism | Random assortment of chromosomes during meiosis. | Active “copy-and-paste” into the wild-type chromosome. |
| Probability of Inheritance | 50% (1 in 2 chance). | Super-Mendelian (91.4% – 99.6%). |
| Population Impact | Traits disappear if they carry a fitness cost. | Drives can outweigh fitness costs to dominate a population. |
The secret to this biased inheritance lies in a specific biological “copy-and-paste” mechanism at the molecular level.
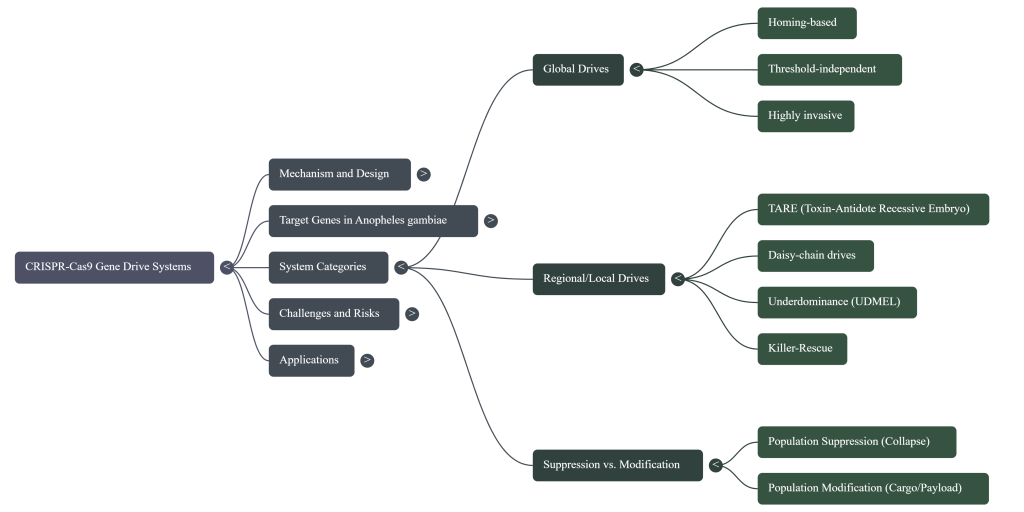
2. Homing Mechanisms and Super-Mendelian Inheritance
Homing drives are biologically inspired by natural selfish genetic elements known as homing endonuclease genes (HEGs). These systems function by converting a heterozygote into a homozygote within the germline, a process termed “homing.” When applied to the malaria vector Anopheles gambiae, these drives target genes essential for female fertility, effectively collapsing reproductive capacity as the drive spreads through the population.

2.1 The Molecular Toolbox: Components of the Drive
To transform a gene into a “selfish” driver, we need three non-negotiable components. In the laboratory, these are often inserted into specific target genes such as AGAP007280 (the Anopheles ortholog of the nudel gene), which is essential for female fertility.
• Cas9 Nuclease (The “Scissors”): An enzyme that cuts DNA. Its precision is governed by its promoter.
◦ So what? The choice of promoter is critical. The vasa2 promoter often causes “leaky” somatic expression (cutting in body cells), which can lower the organism’s health. The nanos promoter is superior because it is germline-specific, ensuring the “scissors” only work where they are needed for inheritance.
• Guide RNA or gRNA (The “GPS”): A short sequence that directs Cas9 to a 20-nucleotide target site.
◦ So what? Without the gRNA, the scissors are blind. Multiplexing (using multiple gRNAs) acts like a backup system, ensuring that if one target site is mutated, others remain available.
• The Cargo (The “Passenger”): This is the payload, such as a malaria-resistance gene.
◦ So what? Think of the drive as a Vehicle and the payload as the Passenger. If the passenger is too “heavy” (has a high fitness cost), the vehicle may slow down, but the super-Mendelian drive mechanism is designed to force that passenger through the population regardless.
Once these tools are inside the germline cell, they initiate a precise sequence of events to modify the genome.
2.2 The Mechanics of Homing: Converting Heterozygotes to Homozygotes
“Homing” is the process of converting a heterozygote (one drive allele, one wild-type) into a homozygote (two drive alleles). This ensures that nearly 100% of gametes carry the drive.
1. Recognition & Cleavage: The gRNA directs Cas9 to create a Double-Strand Break (DSB) on the wild-type chromosome at the exact location opposite the drive.
2. Repair Template Selection: The cell’s repair machinery sees the break and looks for a template to fix it. Because the drive-carrying chromosome is the identical partner, the cell “chooses” it as the template.
3. Homology-Directed Repair (HDR): This is the “Holy Grail” of repair. The cell copies the entire drive sequence into the broken site.
4. Equilibrium vs. Fixation: While a perfect drive would reach fixation (100% of the population), real-world drives with fitness costs often reach an equilibrium frequency, where the drive is present in all individuals but exists in a stable balance with other alleles.
While this process is powerful, nature has a way of “breaking” the system through imperfect repairs.

2.3 Quantifiable Success in Anopheles gambiae
Research targeting somatic female-fertility loci has demonstrated exceptional efficiency in biasing inheritance. The following table details the transmission rates and phenotypic impacts observed at validated target loci:

| Target Locus | Associated Function / Ortholog | Transmission Rate (%) | Phenotypic Result of Homozygosity |
|---|---|---|---|
| AGAP007280 | nudel ortholog (patterning) | 94.4% – 99.6% | Eggs are laid but do not hatch |
| AGAP011377 | Chitin-binding domain | 91.4% – 100.0% | Complete failure to lay eggs |
| AGAP005958 | yellow-g ortholog | 97.9% – 100.0% | Complete failure to lay eggs |
2.3.1 Targeting Fertility
The “So What?” of targeting female-sterility genes hinges on the principle of haplosufficiency. In these systems, heterozygous females (carrying one drive allele) maintain full fertility, allowing the drive to spread silently. However, the disruption of the nudel ortholog (AGAP007280) is particularly efficient for population load management; while homozygous females lay eggs, the resulting failure to hatch ensures the drive continues to impose a reproductive burden without immediately eliminating the females from the mating pool.
While these high transmission rates provide proof-of-principle for rapid modification, the sustainability of first-generation homing drives is frequently undermined by the rapid emergence of molecular resistance.
Based on research findings, two primary strategic pathways define the mission of population engineering:
• Population Modification (Replacement): The rapid spread of a genetic payload—such as an effector gene conferring parasite resistance—through an existing population. The objective is to achieve a stable genomic equilibrium without necessarily impacting population density.
• Population Suppression: The deliberate reduction or collapse of a target population by inducing reproductive loads. This is achieved by targeting essential genes, such as the female-sterility loci identified in Anopheles gambiae: AGAP005958, AGAP011377, and AGAP007280. Notably, AGAP007280 (the nudel ortholog) is currently the only candidate meeting the “minimum requirement” for suppression in modeling due to its milder fertility reduction in heterozygotes compared to the other loci.
3. The Challenge of Molecular Resistance: NHEJ and R-Alleles
Resistance is the primary “Achilles’ heel” for first-generation homing drives. When the drive induces a DSB, the cell may opt for alternative, error-prone repair pathways instead of the desired HDR, creating alleles that the drive can no longer recognize or cut.

3.1 Mechanism of Failure
While HDR is the goal for drive propagation, pathways such as Non-Homologous End Joining (NHEJ) and Microhomology-Mediated End Joining (MMEJ) often intervene. These pathways typically introduce small insertions or deletions (indels) at the target site. These mutations render the protospacer sequence “immune” to the drive’s gRNA, effectively halting its spread.
3.2 R1 vs. R2 Alleles
Resistance alleles are categorized by their impact on the host:
• r1 Alleles (Functional/Resistant): These mutations alter the DNA sequence enough to block the gRNA but preserve the gene’s underlying function. For suppression drives, r1 alleles are catastrophic, as they provide a functional “rescue” that allows the vector to outcompete the drive.
• r2 Alleles (Non-functional/Resistant): These mutations block the gRNA and disrupt gene function. While they do not provide a rescue for the population, they still impede the drive’s ability to “home.”
3.3 Pregonial Resistance
A significant barrier to drive stability is “pregonial resistance.” Research indicates that Cas9 is often active in pregonial cells—mitotically dividing germline cells—before meiosis occurs. This pre-meiotic activity allows a single resistance event to be clonally expanded during germline division, leading to “clusters” of identical resistant offspring from a single parent. This significantly accelerates the rate at which a population becomes immune to the drive construct.
4. Strategic Mitigation: Multiplexing and Essential Gene Recoding
To circumvent these molecular barriers, the field has transitioned toward “evolutionary stable” proposed architectures. These designs shift the strategy from simple disruption to sophisticated recoding and redundant cleavage.
4.1 The Multiplexing Advantage
One of the most robust strategies against resistance is the utilization of multiple gRNAs targeting adjacent sites. Multiplexing requires an organism to evolve resistance at all target sites simultaneously to stop the drive. Crucially, when multiple sites are cleaved, the cell’s repair machinery often results in the loss of the intervening DNA sequence. This process virtually guarantees a non-functional (r2) deletion, preventing the emergence of functional r1 alleles that are common with single-site targets.

4.1.1 Architecture of Multiplexed Guide RNA (gRNA) Systems
gRNA multiplexing provides a critical redundancy layer designed to suppress germline resistance. By targeting multiple adjacent sites, the drive ensures that r1 mutations at a single site do not confer immunity, as remaining wild-type sites remain susceptible to cleavage pressure.
Technical Requirements for gRNA Configuration
1. Multiplexed Target Sites: Constructs must utilize at least two, and ideally up to five, gRNAs. Spacing between target sites must be tailored to the drive mechanism:
◦ Homing Drives: Target sites should be positioned ~100–200 nucleotides (nt) apart to facilitate large deletions that ensure r2 formation during misrepair.
◦ TARE Drives: Target sites must be positioned >1,000 nt downstream from the drive allele. This distance is critical to isolate the cut sites from the recoded rescue template, preventing the cell from using the rescue sequence as a template for incomplete HDR that could generate a functional r1 resistance allele.
2. tRNA Spacer Integration: To facilitate mature gRNA processing from a single Pol III transcript, tRNA spacers must be integrated between individual gRNA sequences, utilizing the host cell’s endogenous RNase P and RNase Z machinery.
3. Promoter Specification: To maintain genomic stability and avoid intramolecular recombination within the gRNA cassette, alternating Pol III promoters (e.g., U6:1 and U6:3) must be utilized for gRNA expression. Note: Cas9 expression must be driven by a distinct germline-specific Pol II promoter (e.g., nanos or vasa); Pol III promoters are strictly reserved for the gRNA cassette.
The strategic advantage of this configuration lies in simultaneous cuts. When multiple gRNAs induce concurrent DSBs, the intervening DNA sequence is typically lost during end-resection. This creates large deletions that effectively guarantee an r2 (non-functional) repair outcome, which the TARE mechanism then exploits for selection.
4.2 Recoding Essential Genes
Advanced mitigation involves targeting the 3′ end of an essential or haploinsufficient gene. The drive construct includes a “genetically recoded” version of the target gene’s sequence to restore functionality upon successful HDR. If the cell attempts repair via NHEJ, the essential gene is disrupted without rescue, leading to the elimination of that non-drive allele from the population. This creates a powerful selective pressure favoring the drive allele.
4.2.1 Design Parameters for the Recoded Antidote
• Codon De-optimization for Homology Reduction: The antidote sequence must utilize synonymous codons optimized for the target organism (e.g., D. melanogaster or An. gambiae). This recoding must maximize sequence divergence from the wild-type target to prevent gRNA cross-reactivity and reduce the likelihood of undesired HDR.
• Homology-Removal Strategy: To prevent “incomplete HDR” that could regenerate an r1 allele, all introns must be eliminated from the rescue sequence. Furthermore, the 3′ UTR of the target gene must be replaced with a non-homologous version (e.g., SV40). This prevents the cell from using the 3′ UTR as a homologous template to copy only the rescue sequence without the full drive construct.
• Target Selection (Essential Genes): Target sites must be located within haplosufficient somatic genes or genes conferring recessive female sterility. Candidate loci include:
◦ AGAP007280: The Anopheles ortholog of the Drosophila nudel gene, essential for dorsoventral patterning.
◦ AGAP011377: A chitin-binding domain gene essential for fertility.
◦ h (hairy): A haplosufficient somatic gene in Drosophila.
The selection pressure of the TARE system ensures that r2/r2 homozygotes—individuals receiving only disrupted alleles—are nonviable. This systematically removes resistance from the gene pool, favoring the drive/antidote construct.
4.3 Somatic vs. Germline Control: Promoters as Strategic Variables
The choice of promoters—specifically nanos versus vasa—is a primary determinant of fitness.
• Superiority of nanos: The nanos promoter is highly germline-specific. It minimizes “leaky” somatic expression, ensuring Cas9 activity is confined to reproductive cells.
• Weakness of vasa: The vasa promoter often exhibits somatic leakage, leading to the formation of resistance alleles in non-reproductive tissues. This somatic nuclease activity can result in significant fitness costs or even accidental sterility in drive-carrying individuals.
Comparison of vasa and nanos Promoters
• vasa Promoter: Often exhibits significant “leaky” somatic Cas9 expression. This leads to high rates of somatic mosaicism, where wild-type alleles are converted to r2 alleles in somatic tissues, imposing severe fitness costs and reducing the reproductive capability of drive-carrying individuals.
• nanos Promoter: The superior choice for high-stability systems. It provides higher germline specificity and minimizes somatic disruption. Species-specific promoter validation is required, as Anopheles and Drosophila exhibit divergent embryo resistance rates under the same regulatory elements.
Carryover Effects and Male Germline Advantage
Maternal carryover occurs when Cas9 protein-gRNA complexes are deposited into the egg, leading to formation of embryonic resistance alleles before HDR can occur. This is a primary driver of r2 allele accumulation. Conversely, an autosomal drive expressed in the male germline (e.g., targeting the cinnabar locus) effectively suppresses embryonic resistance. Because sperm cells are significantly smaller than oocytes, they do not transmit significant quantities of Cas9/gRNA protein into the zygote, preventing the formation of resistance alleles in the paternally inherited genome.
Even with these improvements, the global invasiveness of homing drives necessitates more controllable, threshold-dependent alternatives for localized releases.
Summary: The Cheat Sheet for Overcoming Resistance
| Challenge | Proposed Strategy | Primary Benefit (The “So What?”) |
|---|---|---|
| Somatic Expression | nanos Promoter | Prevents “mosaic” fitness costs by ensuring Cas9 stays out of non-reproductive tissues. |
| r1 Allele Formation | Multiplexing gRNAs | Forces large deletions; converts dangerous “cost-free” resistance into “costly” non-functional resistance. |
| Embryo Resistance | Male Germline Drive | Paternal sperm carry less Cas9 than eggs, preventing NHEJ mutations in the early zygote. |
| Uncontrolled Global Spread | TARE Drive | Creates a threshold requirement; allows for regional, confinable control of genetic modifications. |
5. TARE Drives: A Model for Regionally Confinable Modification
Toxin-Antidote Recessive Embryo (TARE) drives represent a significant evolution in biosafety, providing a mechanism for regionally confined biological control.
5.1 TARE Mechanism
The TARE system operates on a toxin-antidote principle. The drive uses Cas9/gRNA to disrupt a haplosufficient, essential gene (the toxin) across the population, while the drive construct itself carries a recoded, functional version of that gene (the antidote). Unlike homing drives, TARE does not strictly rely on copying itself via HDR; instead, it works by systematically eliminating any offspring that do not inherit the drive allele, as those individuals inherit only disrupted, lethal versions of the target gene.
5.2 Threshold Dynamics and “So What?”
The defining feature of TARE is its threshold-dependent invasion. A TARE drive only spreads if its initial introduction frequency is above a specific, fitness-dependent threshold (e.g., the 24% introduction frequency observed in Drosophila models). In large cage populations, TARE drives have been observed to spread to all individuals in just six generations post-release. If the frequency falls below this threshold—for instance, through low-level migration to a non-target region—the drive is naturally eliminated by selection.
5.3 Same-Site Advantage
Current research distinguishes “same-site” TARE systems from “distant-site” systems like ClvR. The critical advantage of same-site systems is the use of the target gene’s natural promoter elements for the rescue sequence. This maximizes the likelihood of restoring full biological fitness to the insect, avoiding the fitness costs often associated with the artificial promoters used in distant-site architectures.
5.4 Comparative Mechanics: Homing Drives vs. TARE Systems
The architectural selection between homing and toxin-antidote mechanisms dictates the invasion dynamics and geographical confinement of the drive. While standard homing drives prioritize invasiveness, newer architectures prioritize stability and regional control.
Comparison of Drive Architectures
| Feature | Standard Homing Drives | High-Stability TARE Drives |
|---|---|---|
| Mechanism of Action | Super-Mendelian inheritance via HDR-mediated conversion. | Cleavage of wild-type allele; formation of recessive lethal (r2) “Toxin”; rescue via recoded “Antidote.” |
| Susceptibility to Resistance | High; single r1 mutations stop the drive and outcompete it. | Low; r2 alleles are lethal in homozygotes and removed via selection. |
| Invasion Type | Global; invasive at almost any release frequency. | Threshold-dependent; requires minimum frequency to overcome fitness costs. |
| Primary Failure Mode | Formation of functional r1 resistance alleles. | High fitness costs relative to the introduction threshold. |
The “Toxin-Antidote” principle, demonstrated by the “same-site” TARE system at the h (hairy) locus in Drosophila, removes the reliance on narrow temporal windows for HDR. By utilizing the nanos promoter for Cas9 and the U6:3 promoter for gRNAs, the system ensures that embryos with two disrupted alleles are non-viable. Unlike the active conversion of homing drives, TARE drives spread via the selective removal of wild-type alleles. This difference necessitates threshold-based deterministic modeling, providing a mechanism for regional confinement where the drive is lost if migrants fall below the required frequency.
6. Conclusion: Design Principles for Future Gene Drives
Gene drive research has matured from theoretical proofs to stable, engineered systems capable of addressing global crises. The transition from maximizing transmission to prioritizing stability and confinement reflects a sophisticated understanding of population genetics and molecular repair.
Critical Success Factors
Analysis of current source materials identifies three core design principles:
1. Multiplexing: Use multiple gRNAs to ensure that simultaneous cuts lead to the loss of intervening DNA, effectively neutralizing r1 resistance.
2. Strategic Targeting: Target highly deleterious or essential loci to ensure that non-drive repair events are rapidly selected out of the population.
3. Neutral Cargo: Ensure cargo genes for disease resistance are as close to fitness-neutral as possible to prevent the drive from being outcompeted by wild-type alleles.
While these tools offer transformative potential for malaria control, the path to implementation requires disciplined adherence to Arthropod Containment Level 2 (ACL-2) standards. These standards are not merely safety protocols; they are the prerequisite for maintaining public trust and securing the “social license” necessary for eventual regional releases.
Image Summary





Question/Answers
1. How do CRISPR-based gene drives transform traditional methods of agricultural pest population control?
CRISPR-based gene drives transform traditional agricultural pest control by shifting from external interventions, such as recurring chemical sprays, to self-propagating biological mechanisms that manipulate inheritance at the molecular level. Traditional methods, including synthetic insecticides and transgenic Bt crops, often provide only short-term relief and face escalating challenges from field-evolved resistance and environmental toxicity. In contrast, gene drives utilize super-Mendelian inheritance to ensure that a specific genetic trait is passed to nearly 100% of offspring, allowing a desired mutation to spread rapidly through an entire population even if it reduces individual fitness.
Key transformations introduced by this technology include:
• Efficiency of Scale: Unlike the traditional Sterile Insect Technique (SIT), which requires the continuous, massive release of lab-reared sterile males to be effective, gene drives are self-sustaining. In theory, releasing just a few individuals carrying a drive can lead to a complete population invasion within 15 to 20 generations.
• Species-Specific Precision: While traditional pesticides often harm beneficial non-target organisms, CRISPR gene drives are highly specific to the target species because they rely on sexual reproduction for dissemination. This allows for the precise targeting of genes essential for survival or reproduction, such as the doublesex (dsx) gene, to induce female sterility and trigger population collapse.
• Targeted Population Suppression and Alteration: Drives can be designed for suppression, such as “X-shredders” that distort sex ratios in favor of males to eventually cause a population crash, or for alteration, where a population is rendered susceptible to existing pesticides or unable to transmit pathogens.
• Customizable Confinement: Advanced drive architectures offer more control than traditional broad-scale methods. For instance, Daisy-chain drives are designed to be temporally self-limiting and eventually disappear from the population, while Toxin-Antidote Recessive Embryo (TARE) drives exhibit threshold-dependent dynamics that can confine the drive to a specific regional population.
Despite these advantages, the primary technical obstacle remains the evolution of resistance alleles. Resistance occurs when the cell repairs CRISPR-induced double-strand breaks through non-homologous end joining (NHEJ) instead of the desired homology-directed repair, creating mutations that the guide RNA no longer recognizes. To overcome this, researchers are employing gRNA multiplexing—targeting multiple sites within a single gene—so that an insect would have to simultaneously acquire resistance at all sites to stop the drive. By targeting functionally constrained regions of essential genes, any resistant variant that does arise is likely to be non-viable, thereby preventing the spread of resistance and ensuring the drive’s stability in the field.
2. What technical strategies and mechanisms are being developed to prevent environmental gene resistance?
To prevent or mitigate the development of gene drive resistance—primarily caused by non-homologous end joining (NHEJ) or microhomology-mediated end joining (MMEJ) repair that creates “indels” at target sites—researchers are developing several integrated technical strategies and mechanisms.
1. Guide RNA (gRNA) Multiplexing
One of the most promising strategies is gRNA multiplexing, which involves using a single drive construct to target multiple adjacent sites within a gene.
• Mechanism: For resistance to block the drive, an individual would have to simultaneously acquire mutations at all target sites.
• Deletion of Intermediate Sequences: Simultaneous cleavage at multiple sites can result in the loss of intervening DNA, creating a definitive knockout that prevents the formation of “r1” alleles (resistant variants that still maintain gene function).
• Efficiency: Experimental data in Drosophila shows that adding a second gRNA significantly increases drive conversion efficiency while reducing germline resistance rates.
2. Targeting Functionally Constrained and Essential Genes
Resistance can be countered by selecting target sequences where mutations are highly likely to be deleterious or lethal.
• Highly Conserved Loci: Designing drives to target highly conserved regions (e.g., the intron 4–exon 5 boundary of the doublesex gene) ensures that any nuclease-resistant variant arising through NHEJ will likely fail to encode a functional protein, thereby preventing those resistant alleles from being positively selected.
• Essential Gene Targeting: By targeting essential or haplosufficient genes and including a recoded version of that gene in the drive construct to rescue function, any resistant variant that does not inherit the drive will be non-viable and removed from the population.
3. Toxin-Antidote Mechanisms (TARE/ClvR)
New drive architectures, such as Toxin-Antidote Recessive Embryo (TARE) and Cleave and Rescue (ClvR), do not rely on homology-directed repair (HDR) for their initial spread.
• The “Toxin”: The drive construct contains Cas9 and gRNAs that target and disrupt an essential wild-type gene.
• The “Antidote”: The construct also carries a recoded version of the target gene that is functional but cannot be cleaved by the drive’s gRNAs.
• Outcome: This system converts wild-type alleles to disrupted alleles; embryos that inherit two disrupted alleles (and no drive/antidote) die, which systematically increases the frequency of the drive allele without being thwarted by typical NHEJ-mediated resistance.
4. Improved Spatiotemporal Control via Promoters
Resistance rates are often exacerbated by “leaky” somatic expression or maternally deposited Cas9 that acts in the early embryo.
• Germline-Specific Promoters: Utilizing promoters like nanos or zpg (rather than vasa) significantly restricts Cas9 expression to the germline. This reduces somatic mutations that lower fitness and limits the formation of embryo-stage resistance alleles.
• Male Germline Drive: Restricting activity to the male germline can entirely eliminate embryo resistance caused by paternal carryover, as sperm transmit negligible amounts of Cas9 compared to eggs.
5. Molecular Safeguards and Reversal Drives
In cases where a drive must be inactivated or stopped, researchers are developing remediation systems.
• ERACR and CHACR: These elements (Element for the Reversal of the Autocatalytic Chain Reaction and Construct Hitchhiking on the Autocatalytic Chain Reaction) utilize the Cas9 already present in the original drive to home into and inactivate it, effectively removing the undesirable gene drive from the population.
• Immunizing Reversal Drives: These constructs encode their own Cas9 and gRNAs to both remove the original drive and “immunize” wild-type individuals by modifying their target sites to be drive-resistant.
3. How does the intentional release of gene drives impact ecological stability and public interest?
The intentional release of gene drives represents a paradigm shift in biological intervention, offering the potential to permanently eliminate target species or alter wild populations at a global scale. While this technology holds significant promise for humanitarian and economic goals, it also introduces substantial risks to ecological stability and challenges existing frameworks for public interest and ethics.
Impact on Ecological Stability
The primary ecological concern is the unpredictable nature of species removal and the potential for far-reaching environmental disturbances.
• High Invasiveness and Global Spread: Mathematical models indicate that even the least effective “self-propagating” drives are highly invasive; the release of just a few individuals could lead to the infection of an entire species across international borders, making local confinement difficult.
• Trophic Cascades and Niche Replacement: Removing a “pest” species can trigger unintended cascades within an ecosystem, such as the release of mesopredators or the rapid occupation of the vacant niche by another invasive species with similar niche overlap.
• Horizontal Gene Transfer: There is a risk that modified genetic sequences could be transferred to non-target species through interspecific breeding, potentially manifesting deleterious traits in beneficial organisms.
• Environmental Variability: Foundational research is often conducted in static laboratory settings (e.g., 27°C and 75% humidity), which do not reflect the dynamic climatic fluctuations of the real world. Factors like shifting temperatures and humidity can significantly alter mosquito fitness, innate immunity, and the efficacy of the gene drive’s transmission-blocking mechanism.
• Resistance Evolution: The emergence of drive-resistant alleles—often created by the drive’s own error-prone repair mechanisms (NHEJ)—can block the spread of a drive, potentially leading to a resurgence of the pest population with new genetic characteristics.
Impact on Public Interest
The potential for gene drives to address global crises has generated intense interest, but it is accompanied by deep ethical and governance concerns.
• Humanitarian and Economic Benefits: Gene drives could revolutionize public health by eradicating vector-borne diseases like malaria and Zika, and agriculture by reversing insecticide resistance in pests to increase crop yields.
• Ethical Dilemmas of “Wild GMOs”: The technology provides an unprecedented power to “domesticate” almost any sexually reproducing species, challenging the definition of what is “natural” or “wild”. This raises questions about the rights of wild species to survive without human exploitation.
• Public Trust and Social Backlash: Unauthorized or accidental releases could severely damage public trust in science and governance, potentially precluding the future use of beneficial biotechnologies.
• Environmental Justice: Decisions regarding gene drive release are often made in centers far removed from the actual production or release sites, raising concerns that long-term ecological risks will be borne primarily by local human populations.
• Need for Governance: Currently, there is a dearth of regulations specifically tailored to the widespread environmental release of gene drives, necessitating a shift from speculative risk assessment to a systemic approach involving lawmakers and civil society.
To mitigate these impacts, researchers are developing safeguards such as “daisy-chain” drives that are temporally self-limiting and “reversal” drives designed to overwrite and remove engineered genes if unwanted consequences occur. However, even with these tools, the consensus among the sources is that the release of gene drives requires extensive public debate and an abundance of caution due to their potential for irreversible global impact.
4. What are the risks of drives escaping to native ranges?
The escape of gene drives to their native ranges—regions where the target species is an indigenous and often valued part of the ecosystem—poses a significant biological and international biosafety risk. Because many gene drives are designed to be highly invasive and self-propagating, they have the potential to spread far beyond their intended geographic boundaries.
The specific risks and mechanisms associated with drives escaping to native ranges include:
1. High Invasiveness and Global Spread
Threshold-independent drives, such as standard homing gene drives, are considered “global” because they can theoretically invade any population connected by even minimal gene flow. Mathematical models show that the release of just a few engineered individuals can lead to a complete population invasion within 15 to 20 generations. This means that a drive could spread indefinitely through a species unless limited by naturally arising resistance. Consequently, self-propagating drives are generally best suited only for applications intended to affect every wild population of a target species, such as malaria prevention.
2. Mechanisms of Unintended Dispersal
Drive-carrying organisms can move between the target area and native ranges through several pathways:
• Anthropogenic Movement: Invasive species are frequently associated with human trade and movement, which acts as a “stepping stone” between populations.
• Cargo Stowaways: Commensal species like house mice and black rats are particularly prone to escaping through biosecurity pathways by hitchhiking on shipping vessels or aircraft.
• Natural Dispersal: Even in ostensibly isolated areas like islands, there is a high likelihood of unwanted spread across international borders through natural migration or weather-driven events.
3. Vulnerability of Native Populations
While a gene drive might be deployed to suppress a pest population in one region, the native populations in their original range remain highly susceptible to the deleterious traits. For instance, a drive designed for population suppression through female sterility could return to the species’ home range and cause a total population collapse or even global extinction. This represents a severe threat to global biodiversity, especially if the target species is a keystone species in its native environment.
4. Categorical Risk by Drive Architecture
The level of escape risk is closely tied to the design of the drive:
• Global/Threshold-Independent Drives: These have the highest risk as they can spread from very low initial frequencies and survive long-distance dispersal of a few individuals.
• Regional/Threshold-Dependent Drives: Architectures like TARE (Toxin-Antidote Recessive Embryo) or chromosomal translocations are designed for confinement. They must exceed a specific population frequency to spread, making it much less likely for a few rare migrants to establish a drive in a distant native range.
5. International and Social Consequences
Any unauthorized release affecting a wild population in its native range could severely damage public trust in scientists and regulatory bodies. Such events could trigger substantial social backlash, potentially precluding future beneficial applications of biotechnology. Furthermore, the lack of international agreements and current regulatory gaps for “wild GMOs” complicates the management of these cross-border biosecurity risks.
5. What are the risks of using recoded versions of essential genes?
Using recoded versions of essential genes—typically as part of Toxin-Antidote Recessive Embryo (TARE) or homing-based drive architectures—is intended to reduce the spread of resistance by ensuring that any non-functional mutation results in death. However, this strategy introduces several significant technical and ecological risks.
1. Formation of Functional Resistance Alleles (r1 Alleles)
The most critical technical risk is the potential for the drive to generate r1 resistance alleles, which are mutations that preserve the function of the target gene but are “immune” to the drive’s guide RNAs.
• Undesired Homology-Directed Repair (HDR): In systems containing a recoded rescue element, there is a risk that the cell’s repair machinery will use the recoded portion as a template to repair a break without copying the entire drive construct. This results in an allele that provides the necessary “rescue” for survival but does not carry the drive or its payload, allowing these alleles to outcompete the drive construct.
• Incomplete HDR: Even if the system is designed to minimize homology, incomplete HDR events can occur where the target gene is repaired (potentially using the recoded sequence) without successfully inserting the full drive, thereby creating a viable resistance allele.
2. Fitness Costs and Somatic Leakiness
While recoding aims to restore the function of an essential gene, the process itself can negatively impact the organism’s viability.
• Leaky Somatic Expression: If the nuclease (Cas9) is expressed in somatic cells rather than being strictly restricted to the germline, it may cleave the remaining wild-type allele in those tissues. In an individual carrying only one functional recoded gene, this somatic disruption can lower fitness or cause mortality, even if the germline is successfully modified.
• Suboptimal Recoding: Although studies in E. coli and yeast suggest that essential genes can often be recoded with minimal impact, there is always a risk that the synthetic sequence will alter RNA-binding motifs, secondary structures, or codon usage in a way that reduces the overall fitness of the modified organism compared to wild-type individuals.
3. Loss of Drive Confinement
Ecological risks arise if the drive construct becomes unstable over time or generations.
• Payload Loss: If a gene drive carries a deleterious payload (such as one for population suppression) alongside a recoded rescue gene, there is a risk that the payload could be lost while the recoded “antidote” remains.
• Unintended Global Spread: Because the “antidote” (the recoded gene) reduces the fitness cost of carrying the drive, a drive that has lost its payload becomes less deleterious and could transition from a “regional” drive to a highly invasive “global” drive, spreading far beyond intended geographic boundaries.
4. Technical Constraints in Design
Designing these systems requires a difficult balance of genomic distances. To prevent undesired HDR, researchers must place the drive’s cut sites a large distance from the recoded rescue element (often several hundred to 1,400 nucleotides away). If these elements are too close, the rate of functional resistance allele formation increases; if they are too far, the efficiency of the initial drive conversion may be compromised because the homology arms may no longer effectively facilitate the insertion of the large drive cassette.
6. How do Toxin-Antidote systems differ from traditional homing drives?
Toxin-Antidote (TA) systems, such as Medea and Toxin-Antidote Recessive Embryo (TARE) drives, differ from traditional homing drives primarily in their mechanism of propagation, their level of invasiveness, and their vulnerability to resistance alleles.
1. Mechanism of Propagation
The fundamental distinction lies in how these drives increase their frequency within a population:
• Traditional Homing Drives: These utilize a “cut-and-copy” mechanism. The drive construct encodes an endonuclease (like Cas9) that creates a double-strand break (DSB) at a specific target site on the homologous wild-type chromosome. If the cell repairs this break using homology-directed repair (HDR), the drive allele is copied into the target site, converting a heterozygote into a homozygote.
• Toxin-Antidote Systems: These do not rely on active copying or HDR for their initial spread. Instead, they function by eliminating individuals that do not inherit the drive. In a TARE drive, the “toxin” is the disruption of an essential gene (creating recessive lethal alleles), while the drive construct carries a recoded “antidote” version of that gene to rescue its carriers. The drive spreads because offspring that inherit two disrupted alleles without a drive/antidote allele are nonviable and removed from the population.
2. Invasiveness and Confinement (Threshold Dynamics)
The two systems exhibit vastly different population dynamics, categorized as “global” versus “regional” spread:
• Homing Drives (Global): These are considered threshold-independent, meaning that in deterministic models, they are self-sustaining and can theoretically invade an entire population starting from just a single individual. This high invasiveness makes them difficult to confine to specific geographic regions.
• TA Systems (Regional): Systems like TARE and Medea are threshold-dependent. They only spread if they are introduced into a population above a specific frequency (determined by the fitness costs of the drive). If the frequency remains below this threshold—for instance, due to rare long-distance migration—the drive will be eliminated by selection, effectively confining the drive to a target region.
3. Vulnerability to Resistance
Resistance alleles, often created by the drive’s own cleavage activity via non-homologous end joining (NHEJ), impact the two systems differently:
• Homing Drives: These are highly sensitive to resistance. If an error-prone repair creates a mutation (an “r1” or “r2” allele) that the drive’s guide RNA can no longer recognize, that allele will have a fitness advantage over the costly drive and can block its spread.
• TARE Drives: These are significantly more resilient to resistance. Because TARE uses the creation of disrupted, lethal alleles to drive itself, the formation of “r2” (loss-of-function) resistance alleles by NHEJ actually aids the drive by contributing to the pool of nonviable genotypes. The only major threat is the formation of “r1” (functional) resistance alleles, which can be minimized by targeting essential genes with multiple gRNAs.
4. Promoter and Temporal Requirements
• Homing Drives: These require strict germline-specific promoters (e.g., nanos or zpg) to ensure cleavage occurs during a narrow window where HDR is the preferred repair pathway. Embryonic activity or maternal carryover of Cas9 is problematic because it creates resistance alleles that prevent further homing.
• TARE Systems: These offer greater flexibility. While germline activity is used, maternal deposition of Cas9 and gRNA in the embryo is actually desirable, as it ensures that wild-type alleles are disrupted in the next generation, thereby accelerating the drive’s spread.
7. What happens if a drive loses its payload but keeps the antidote?
If a gene drive, particularly a Toxin-Antidote (TARE) system, loses its payload but retains the antidote (the recoded rescue element), the primary result is a significant reduction in the fitness cost associated with the drive, which can lead to unintended global spread.
The following consequences and mechanisms are associated with this scenario:
Increased Invasiveness and Loss of Confinement
• Threshold Shift: TARE drives are designed to be “regional” because they exhibit threshold-dependent dynamics. They only spread through a population if they are introduced above a specific frequency, which is determined by the drive’s fitness cost.
• Global Spread: If the fitness costs of a drive are primarily caused by its payload, losing that payload removes the biological “brake” that keeps the drive confined to a specific region.
• Conversion to “Global” Drive: A drive that has lost its payload may see its introduction threshold drop toward zero in idealized models. This allows the drive—which still carries the nuclease and the antidote—to become highly invasive, potentially spreading across international borders to a much larger area than the payload was intended to reach.
Functional Persistence Without Phenotypic Effect
• Continued Propagation: Even without the payload, the drive remains a “selfish” genetic element. The nuclease will continue to cleave wild-type alleles, and the antidote will continue to rescue those who inherit the drive.
• Ineffectual Modification: The drive will successfully propagate and replace wild-type alleles with its own sequence, but it will fail to provide the desired impact (such as disease resistance or population suppression) because the active “cargo” is missing.
Mechanisms of Payload Loss
• Incomplete Homology-Directed Repair (HDR): This scenario often occurs during the “homing” process if the cell’s repair machinery uses the recoded rescue element as a template but fails to copy the entire large drive cassette, including the payload.
• Genetic Instability: Large drive constructs are more susceptible to incomplete HDR. To mitigate this risk, researchers attempt to place target sites far from the recoded rescue elements to ensure the drive is copied as a single, stable unit.
In summary, losing the payload while keeping the antidote results in a “fit” but empty drive that can infect entire species globally while failing to achieve its humanitarian or agricultural goal.
8. How do fitness costs influence the speed of payload loss?
Fitness costs associated with a payload significantly influence the speed and dynamics of payload loss by creating selective pressure that favors variants which have discarded the costly genetic material. In systems like the Toxin-Antidote Recessive Embryo (TARE) drive, payload loss typically occurs through undesired homology-directed repair (HDR), where the cell’s repair machinery copies the recoded rescue element (the “antidote”) but fails to include the large, often deleterious payload sequence.
The influence of fitness costs on the speed of this process can be understood through several key mechanisms:
• Selective Advantage of Deletion Variants: Genetic payloads are often designed to impose high fitness costs to achieve population suppression or to carry heavy metabolic loads for population modification. When a variant arises that has lost this payload through incomplete HDR, it immediately gains a substantial selective advantage over the full drive construct because it retains the “fit” antidote but lacks the “costly” cargo. The greater the fitness cost of the original payload, the faster this “fit” but empty drive will outcompete and replace the full drive in the population.
• Shift in Invasiveness Thresholds: High fitness costs from a payload create a nonzero introduction threshold, which is the biological “brake” that allows a drive to be “regional” or confined to a specific area. If the payload is lost, this cost-induced threshold drops significantly, potentially converting a regional drive into a highly invasive “global” drive. This transition allows the payload-less drive to spread much more rapidly and over a larger geographic area than the original construct was intended to reach.
• Impact on Spread and Fixation: High fitness costs can prevent a drive from ever reaching fixation, causing it instead to reach a lower equilibrium frequency. In these scenarios, the drive remains in a prolonged state of flux where it is more vulnerable to being outpaced by more fit alleles, such as those that have undergone payload loss or have acquired functional resistance.
• Evolutionary Instability: Mathematical models indicate that even if a drive is initially successful, resistance alleles or deletion variants that confer no fitness cost to the host will inevitably spread at the expense of a costly drive. High payload costs accelerate this instability, as the selective gap between the drive and the “escaped” variant is wider, leading to a faster collapse of the drive’s effectiveness.
To mitigate these risks and slow the speed of payload loss, researchers emphasize minimizing homology between the cut sites and the rescue element and selecting target sites located at a large distance from the drive insertion site to ensure the construct is copied as a single, stable unit.
9. What happens if a drive outcompetes the original construct?
If a variant—such as a resistance allele or a drive construct that has lost its payload—outcompetes the original gene drive construct, the primary result is the evolutionary failure of the intervention’s goal, whether that be population suppression or modification. This occurs because natural selection favors any variant that carries a lower fitness cost than the original engineered element.
The following specific outcomes occur when a drive is outcompeted:
1. Shift from Super-Mendelian Spread to Natural Selection
While a gene drive spreads rapidly through a population via super-Mendelian inheritance (often reaching high frequencies in 15–30 generations), its replacement by a more “fit” variant is driven by natural selection, which is typically a much slower process. For example, a resistance allele might take more than 100 generations to reach 50% frequency, even if the drive it is replacing achieved 99% frequency in less than 30 generations.
2. Failure of Population Suppression
In suppression strategies, such as those targeting the doublesex (dsx) gene or other female-fertility genes, the emergence of a functional resistance allele (r1) is the main technical obstacle.
• If an r1 allele arises—typically through error-prone non-homologous end joining (NHEJ)—it restores the function of the target gene while remaining “immune” to the drive’s nuclease.
• Because individuals carrying the r1 allele are fertile and do not suffer the heavy fitness costs of the suppression drive, they will eventually replace the drive, allowing the pest population to rebound to its original levels.
3. Loss of the Desired Payload (Population Modification)
In modification strategies, the drive is designed to carry a “payload,” such as a gene rendering mosquitoes resistant to malaria.
• If the drive construct undergoes incomplete homology-directed repair (HDR), it may copy the drive mechanism but fail to include the large payload sequence.
• If the payload imposes a fitness cost, this “empty” drive variant will have a substantial selective advantage over the original construct.
• The result is a population fixed for the drive mechanism (nuclease and gRNA) but lacking the therapeutic trait, rendering the intervention useless.
4. Conversion to an Invasive “Global” Drive
In Toxin-Antidote (TARE) systems designed for regional confinement, outcompeting by a payload-less variant can lead to an ecological risk.
• Regional confinement relies on threshold-dependent dynamics, where a high fitness cost (often from the payload) prevents the drive from spreading via rare migrants.
• If the drive loses its costly payload but retains the recoded “antidote” gene, the fitness cost drops.
• This reduction in cost can lower the introduction threshold toward zero, effectively converting a once-controllable regional drive into a highly invasive global drive that can infect the entire species across international borders without achieving its management goal.
5. Inevitability in Large Populations
Mathematical modeling suggests that drive constructs are inherently unstable in sufficiently large populations. If a fully resistant, cost-free allele (Sn) has any nonzero rate of production, it will certainly invade and eventually replace a resident drive population that has any associated fitness cost. To mitigate this, researchers are developing strategies like gRNA multiplexing and targeting functionally constrained essential genes to ensure that any variant that outcompetes the drive is likely to be non-viable.
10. What are the specific technical ways to stabilize a payload?
Specific technical ways to stabilize a payload within a CRISPR-based gene drive primarily focus on overcoming the evolution of resistance and preventing the selective advantage of alleles that have lost the payload. Because payloads often impose a fitness cost, they are prone to being outcompeted by “fit” resistance alleles or drive variants that have undergone incomplete homology-directed repair (HDR).
Researchers utilize the following strategies to ensure payload stability:
1. Guide RNA (gRNA) Multiplexing
Using multiple gRNAs to target adjacent sites within a single gene is a fundamental strategy for stability.
• Preventing Functional Resistance: For a functional resistance allele (r1) to block the drive, mutations must occur simultaneously at every target site, which is statistically unlikely.
• Inducing Definitive Knockouts: If cutting occurs at two or more sites simultaneously, the intervening DNA sequence is lost, resulting in a large deletion that ensures the target gene is non-functional, thereby selecting against those resistant variants.
• Efficiency Gains: Experimental data indicates that multiplexing significantly increases germline drive conversion and reduces the formation of resistance alleles compared to single-gRNA systems.
2. Targeting Functionally Constrained and Essential Genes
Stability is enhanced by choosing target sites where mutations are highly likely to be deleterious.
• Selection Against Resistance: By targeting an essential or haploinsufficient gene, any resistance allele that disrupts the gene (r2) will be removed by natural selection.
• Functional Rescue (Recoding): The drive construct can include a genetically recoded version of the essential gene that restores function but is “immune” to the drive’s own gRNAs. This ensures that only individuals who successfully inherit the drive (and its payload) remain viable.
• Targeting Conserved Loci: Targeting highly conserved sequences, such as the doublesex (dsx) intron 4–exon 5 boundary, minimizes the risk of resistance because these regions have rigid functional requirements across species.
3. Optimized Genomic Architecture
The physical arrangement of the drive components is critical to prevent the payload from being “lopped off” during repair.
• Increasing Homology Distance: To reduce the rate of undesired HDR—where the cell copies the recoded rescue element but not the payload—researchers place target sites a large distance (e.g., several hundred to 1,400 nucleotides) from the repair templates.
• Minimizing Homology: Replacing natural 3′UTRs with substitute or recoded sequences can remove homology between the cut sites and the drive components, forcing the machinery to copy the entire drive cassette as a single stable unit.
4. Toxin-Antidote Mechanisms (TARE/ClvR)
Architecture like the Toxin-Antidote Recessive Embryo (TARE) drive stabilizes payloads by changing the spread dynamic.
• Threshold-Dependent Dynamics: These systems spread by rendering wild-type alleles non-viable (the “toxin”) and providing a recoded rescue (the “antidote”), making them more resilient to resistance than traditional homing drives.
• Same-Site Integration: Placing the TARE drive at the same locus as the target gene allows the use of natural promoter elements, which enhances the reliability of the rescue function and reduces the metabolic burden of the construct.
5. Promoter Selection and Germline Restriction
Restricting the nuclease activity to specific tissues and times prevents “leaky” expression that destabilizes the drive.
• Germline-Specific Promoters: Using promoters like nanos or zpg (rather than vasa) restricts Cas9 expression to the germline, preventing somatic mutations that could lower the fitness of carriers or cause premature resistance allele formation in pregonial cells.
• Mitigating Paternal/Maternal Carryover: Stabilizing a drive requires preventing the carryover of active Cas9 into the early embryo, where it can create resistance alleles that bypass the homing process; for example, male germline drives avoid the heavy maternal deposition of Cas9 found in eggs.
11. How does higher fitness cost accelerate the loss of cargo?
Higher fitness costs accelerate the loss of genetic cargo by creating intense selective pressure that favors any variant that manages to retain the drive’s survival mechanism (the antidote or rescue element) while discarding the deleterious payload. Because many gene drives carry a “cargo” gene designed to suppress a population or modify a trait, this cargo often imposes a metabolic or physiological burden that reduces the organism’s overall fitness.
The mechanisms by which higher fitness costs drive this loss include:
1. Selective Advantage of Cargo-less Variants
If a drive construct has a fitness cost, any naturally arising variant that is resistant to the drive’s nuclease but lacks the costly cargo—such as a functional resistance allele (r1)—will have a substantial selective advantage. In a sufficiently large population, if such a cost-free resistant allele has even a minimal rate of production, it is mathematically predicted to certainly invade and eventually replace the costly resident drive population. The greater the fitness cost of the original cargo, the wider the selective gap, and the faster this replacement occurs through natural selection.
2. Incomplete Homology-Directed Repair (HDR)
Loss of cargo often occurs during the molecular “homing” process. In architectures like the Toxin-Antidote Recessive Embryo (TARE) drive, there is a technical risk of undesired HDR. This happens when the cell’s repair machinery uses the drive’s recoded “antidote” as a template but fails to copy the entire large cassette, effectively “lopping off” the cargo.
• Speed of Invasion: Variants that lose the cargo via this mechanism immediately become more “fit” than the full construct.
• Experimental Observations: In simulations of population alteration drives, constructs with even moderate fitness costs (e.g., above 8%) were shown to be highly unstable, often falling below their initial release frequency in fewer than 200 generations as they were outcompeted by more fit alleles.
3. Disappearance vs. Equilibrium
In some scenarios, if the fitness cost of the cargo is high enough to outweigh the drive’s inheritance bias (homing rate), the construct may fail to spread entirely and disappear from the population over time. For example, researchers observed that fitness costs in certain mosquito targets (like AGAP011377) were so high that they effectively acted as a “dominant lethal” system, precluding the drive from reaching fixation.
4. Loss of Regional Confinement
In threshold-dependent “regional” drives, the fitness cost of the cargo acts as a biological “brake” that prevents the drive from spreading to distant populations via rare migrants. However, if the drive loses its costly cargo but keeps its survival antidote:
• The introduction threshold drops significantly, sometimes toward zero.
• The drive can transition from a controllable regional intervention into a highly invasive “global” drive, spreading the nuclease and antidote to a much larger geographic area than the intended cargo ever reached.
To mitigate these risks, researchers suggest that cargo genes should be designed to be as close to neutral as possible to minimize the selective advantage of deletion variants.
12. What genomic design techniques best prevent incomplete homology-directed repair?
Genomic design techniques to prevent incomplete homology-directed repair (HDR) focus on manipulating the distance and homology between the drive’s components and its target sites to ensure the entire construct is copied as a single, stable unit. Incomplete HDR occurs when the cell’s repair machinery uses only a portion of the drive—such as a recoded rescue element—as a template, resulting in functional resistance alleles (r1) that block the drive.
The primary technical strategies to mitigate this risk include:
1. Increasing Genomic Distance Between Cut Sites and Templates
A highly effective method for reducing undesired or incomplete HDR is to place the drive’s cut sites a large distance from the homology templates.
• Distance Dynamics: Research indicates that HDR efficiency decreases by approximately 15% when the distance between cut sites and homology templates reaches ~100 nucleotides on one side. For larger distances of 1,000 nucleotides or more, the rate of incomplete repair falls by over 50%.
• Preferred Templates: By selecting target sites far downstream from the drive insertion site (e.g., ~1,400 nucleotides), the cell’s repair machinery is more likely to use the immediate homology of the original target gene DNA downstream of the drive, rather than reaching across the large cassette to copy only the recoded portion.
2. Eliminating Internal Sequence Homology
Removing all regions of homology between the cut sites and the internal drive components prevents the repair machinery from identifying internal segments as viable templates.
• 3′UTR Replacement: One specific technique is to replace the target gene’s native 3′ untranslated region (3′UTR) with an equivalent but non-homologous sequence. This ensures that the drive cassette is identified by the cell as a single unit that must be copied in its entirety.
• Recoded or Substitute Sequences: If the native 3′UTR must be used in a recoded rescue element, designers should use a smaller 3′UTR or increase the distance between the target site and the 3′UTR. Using substitute or synthetic 3′UTRs is generally preferred to minimize undesired templates.
3. Structural Rearrangement and Recoding
• Rearranging Recoded Regions: Adjusting the physical orientation of recoded segments within the drive construct can further minimize the likelihood of partial templates being utilized during the repair process.
• Comprehensive Recoding: In “proposed” drive architectures, the 3′ end of an essential target gene is completely recoded to restore function while remaining resistant to the drive’s guide RNAs (gRNAs). This architecture selects against any incomplete repair that results in a gene knockout.
4. Mitigating Intramolecular Recombination
Incomplete homing can sometimes be resolved using the homology inherent in the gRNA sequence itself and its cognate target in the genome. This is often the result of intramolecular recombination between regions of homology within the gRNA construct and the endogenous target at the insertion site. Designing constructs with minimal internal homology—such as avoiding repeated promoter sequences that can cause genomic instability—helps prevent these rare non-functional homing events.
13. How can payload loss convert regional drives into global ones?
The conversion of a regional gene drive into a global one through payload loss is a result of the fundamental relationship between a drive’s fitness cost and its invasion threshold.
The Mechanism of Regional Confinement
Regional drives, such as the Toxin-Antidote Recessive Embryo (TARE) drive, are characterized by threshold-dependent dynamics. These systems only spread if they are introduced into a population above a specific frequency. This invasion threshold acts as a biological “brake,” ensuring that the drive remains confined to a target region and cannot establish itself in distant populations via rare migrants. The height of this threshold is directly determined by the fitness cost the drive imposes on the host organism.
The Role of the Payload in Stability
In most population modification strategies, a significant portion of the drive’s fitness cost is caused by the payload allele—the specific gene sequence (such as a malaria-blocking transgene) intended to produce a desired effect. As long as this costly payload remains attached to the drive mechanism, the nonzero introduction threshold persists, maintaining the drive’s status as a “regional” system.
How Payload Loss Triggers Conversion
The transition from a regional to a global drive occurs through a technical failure during the molecular repair process:
• Undesired Homology-Directed Repair (HDR): During the “homing” process, the cell’s repair machinery may use the drive’s recoded rescue element (the antidote) as a template but fail to include the entire large cassette containing the payload.
• Formation of “Fit” Variants: This incomplete repair results in an “empty” drive variant that retains the ability to survive (via the antidote) but lacks the costly cargo.
• Threshold Collapse: Because this variant lacks the fitness cost of the payload, it gains a substantial selective advantage over the original construct. As the fitness cost associated with the drive decreases, the introduction threshold drops toward zero.
• Global Spread: Once the threshold is removed, the drive transitions into a highly invasive “global” system. These global drives are self-sustaining at any frequency and can theoretically invade any population connected by even minimal gene flow.
Consequently, a drive intended for local or regional application can escape its designated boundaries and spread indefinitely through an entire species, carrying the drive components (Cas9 and gRNA) to a much larger area than the intended payload ever reached. This results in a population fixed for the drive mechanism but lacking the phenotypic effect needed for pest control or disease prevention. To mitigate this, researchers attempt to minimize regions of homology and select target sites a large distance from the drive insertion site to ensure the entire construct is copied as a single, stable unit.
14. How does 3′ UTR replacement help ensure drive cassette stability?
Replacing the 3′ untranslated region (UTR) of a target gene with a non-homologous or substitute sequence is a critical technique used to remove all internal homology between the drive’s cut sites and its internal components. In standard architectures that utilize a recoded rescue element to restore gene function, the presence of a native 3′ UTR sequence within the drive can act as an unintended template for homology-directed repair (HDR). If the cell’s repair machinery utilizes this internal 3′ UTR as a template, it can lead to incomplete HDR, where only the recoded “antidote” or rescue portion is copied while the rest of the drive cassette—including the nuclease and payload—is discarded.
This partial repair process creates functional resistance alleles (r1 alleles) that restore the target gene’s function but remain “immune” to further cutting, allowing these “fit” variants to rapidly outcompete the full drive construct and block its spread. By replacing the native 3′ UTR with an equivalent but non-homologous sequence, designers ensure that the entire drive cassette is identified as a single unit for copying. This structural modification minimizes the rate of undesired HDR and reduces the technical requirement of placing target sites a large distance from the drive insertion site to prevent partial template usage. Consequently, 3′ UTR replacement serves as a vital safeguard to ensure the evolutionary stability of population modification and suppression drives by selecting against the formation of viable, cost-free resistant variants.
15. How do climate fluctuations affect gene drive stability?
Climate fluctuations significantly affect the stability and efficacy of CRISPR-based gene drives by impacting the molecular functionality of the drive components, the host’s biological fitness, and the evolutionary dynamics of resistance. Foundational gene drive research is typically conducted under static laboratory conditions (standardized at 27°C and 75% relative humidity), which creates a “lab-to-field” discrepancy because these settings do not reflect the dynamic fluctuations mosquitoes and other pests encounter in nature.
Key ways climate fluctuations affect gene drive stability include:
• Molecular Component Sensitivity: Temperature variations can directly influence the enzymatic activity of the Cas9 nuclease and the stability of the guide RNA (gRNA), which may compromise the super-Mendelian inheritance bias essential for the drive to spread.
• Host Fitness and Spread Efficiency: The successful spread of a gene drive is intrinsically linked to the overall fitness of engineered organisms—including their survival, size, and reproductive success—all of which are highly sensitive to environmental pressures and extreme weather events like heatwaves, floods, or droughts.
• Fluctuating Resistance Probabilities: Climate-driven seasonal changes cause dramatic fluctuations in population sizes (e.g., winter population crashes followed by warm-season booms), which significantly alter the supply of resistance alleles and the probability that these alleles establish in a population. Resistance probabilities may even depend on the specific time of year a driver is released, as alleles arising during warm-season population expansions are more likely to persist through subsequent collapses.
• Altered Promoter Activity: Diurnal environmental variations in temperature and humidity significantly impact the activity of promoters used to drive transgene expression. If a promoter’s precision is compromised by environmental stress, it could lead to suboptimal expression of the drive or unintended somatic activity, both of which reduce the stability of the intervention.
• Temporal Mismatch with Parasites: Ambient temperature shifts can alter the extrinsic incubation period of pathogens (the time required for a parasite to become transmissible within the vector), potentially creating a mismatch between the intensity of a pathogen challenge and the expression window of the drive’s anti-parasitic payload.
• Complex Abiotic Interplay: In natural settings, mosquitoes face a complex interplay of abiotic factors beyond temperature—including precipitation, hydrology, and salinity—that regulate their physiology and vectorial capacity in ways that are not yet fully understood or captured in laboratory models.
Because of these factors, research suggests that the realistically predicted efficacy of a gene drive in a climatically changing world may differ substantially from the performance observed in optimized laboratory settings.
References
Bharti, N., Modi, U., Bhatia, D. D., & Solanki, R. (2026). Engineering delivery platforms for CRISPR-Cas and their applications in healthcare, agriculture and beyond. Nanoscale Advances. https://doi.org/10.1039/D5NA00535
Champer, J., Lee, E., Yang, E., Liu, C., Clark, A. G., & Messer, P. W. (2020). A toxin-antidote CRISPR gene drive system for regional population modification. Nature Communications, 11(1). https://doi.org/10.1038/s41467-020-14960-3
Champer, J., Liu, J., Oh, S. Y., Reeves, R., Luthra, A., Oakes, N., Clark, A. G., & Messer, P. W. (2018). Reducing resistance allele formation in CRISPR gene drive. Proceedings of the National Academy of Sciences of the United States of America, 115(21), 5522–5527. https://doi.org/10.1073/pnas.1720354115
Courtier‐Orgogozo, V., Morizot, B., & Boëte, C. (2017). Agricultural pest control with CRISPR ‐based gene drive: time for public debate . EMBO Reports, 18(6), 878–880. https://doi.org/10.15252/embr.201744205
Dael, L., & Simões, M. L. (2026). Beyond the static lab: environmental variability in genetically modified mosquito target gene identification for malaria control. Current Opinion in Insect Science, 101489. https://doi.org/10.1016/j.cois.2026.101489
Hammond, A., Galizi, R., Kyrou, K., Simoni, A., Siniscalchi, C., Katsanos, D., Gribble, M., Baker, D., Marois, E., Russell, S., Burt, A., Windbichler, N., Crisanti, A., & Nolan, T. (2016). A CRISPR-Cas9 gene drive system targeting female reproduction in the malaria mosquito vector Anopheles gambiae. Nature Biotechnology, 34(1), 78–83. https://doi.org/10.1038/nbt.3439
Kyrou, K., Hammond, A. M., Galizi, R., Kranjc, N., Burt, A., Beaghton, A. K., Nolan, T., & Crisanti, A. (2018). A CRISPR–Cas9 gene drive targeting doublesex causes complete population suppression in caged Anopheles gambiae mosquitoes. Nature Biotechnology, 36(11), 1062–1071. https://doi.org/10.1038/nbt.4245
Malik, D. J., Sokolov, I. J., Vinner, G. K., Mancuso, F., Cinquerrui, S., Vladisavljevic, G. T., Clokie, M. R. J., Garton, N. J., Stapley, A. G. F., & Kirpichnikova, A. (2017). Formulation, stabilisation and encapsulation of bacteriophage for phage therapy. In Advances in Colloid and Interface Science (Vol. 249, pp. 100–133). Elsevier B.V. https://doi.org/10.1016/j.cis.2017.05.014
Marshall, J. M., & Akbari, O. S. (2018). Can CRISPR-Based Gene Drive Be Confined in the Wild? A Question for Molecular and Population Biology. In ACS Chemical Biology (Vol. 13, Number 2, pp. 424–430). American Chemical Society. https://doi.org/10.1021/acschembio.7b00923
Moro, D., Byrne, M., Kennedy, M., Campbell, S., & Tizard, M. (2018). Identifying knowledge gaps for gene drive research to control invasive animal species: The next CRISPR step. In Global Ecology and Conservation (Vol. 13). Elsevier B.V. https://doi.org/10.1016/j.gecco.2017.e00363
Noble, C., Adlam, B., Church, G., Esvelt, K. M., & Nowak, M. A. (2018). Current CRISPR gene drive systems are likely to be highly invasive in wild populations. ELIFE, 7. https://doi.org/10.7554/eLife.33423.001
Noble, C., Olejarz, J., Esvelt, K. M., Church, G. M., & Nowak, M. A. (2017). Evolutionary dynamics of CRISPR gene drives. Science Advances, 3. DOI: 10.1126/sciadv.1601964
Saini, A., Sharma, Neha, Sharma, Nidhi, Kumari, N., Sharma, M., Singh, B., & Thakur, A. K. (2026). Precision pest management: Genome editing tools, specifically CRISPR/Cas9 and future prospects. In Pesticide Biochemistry and Physiology (Vol. 218). Academic Press Inc. https://doi.org/10.1016/j.pestbp.2026.106941
Salaudeen, J. O., Ajibola Iyiola, O., Momoh, F. K., Ngutor, J. A., Simon, W. N., & Hameed, S. (2026). Exploring Anopheles gambiae Giles, 1902, (Family: Culicidae) Doublesex Gene Editing via CRISPR and Its Implications for CAR T-Cell Therapy and Vector Control in Nigeria. World News of Natural Sciences, 64. www.worldnewsnaturalsciences.com
Unckless, R. L., Clark, A. G., & Messer, P. W. (2017). Evolution of resistance against CRISPR/Cas9 gene drive. Genetics, 205(2), 827–841. https://doi.org/10.1534/genetics.116.197285
Webber, B. L., Raghu, S., & Edwards, O. R. (2015). Opinion: Is CRISPR-based gene drive a biocontrol silver bullet or global conservation threat? Proceedings of the National Academy of Sciences of the United States of America, 112(34), 10565–10567. https://doi.org/10.1073/pnas.1514258112
Related posts:
 Traditional Sardinian Fermented Milk Boosts Gut Health by Reshaping Microbiota
Traditional Sardinian Fermented Milk Boosts Gut Health by Reshaping Microbiota
 Boosting Sunflower Growth in Saline Soils: The Role of Phosphate-Solubilizing Bacteria and Phosphorus-Enriched Biochar
Boosting Sunflower Growth in Saline Soils: The Role of Phosphate-Solubilizing Bacteria and Phosphorus-Enriched Biochar
 Broad-Host-Range Phages: Nature’s Ubiquitous Virus Hunters Across Ecosystems
Broad-Host-Range Phages: Nature’s Ubiquitous Virus Hunters Across Ecosystems
 Revolutionizing TUDCA Production: How Engineered Bacillus subtilis Unlocks a Cost-Effective Bear Bile Alternative
Revolutionizing TUDCA Production: How Engineered Bacillus subtilis Unlocks a Cost-Effective Bear Bile Alternative
 Bacteriocins vs. Biofilms: A Low-Cost Breakthrough in the Fight Against Antibiotic Resistance
Bacteriocins vs. Biofilms: A Low-Cost Breakthrough in the Fight Against Antibiotic Resistance