1. Introduction: The Strategic Imperative for Speed Breeding
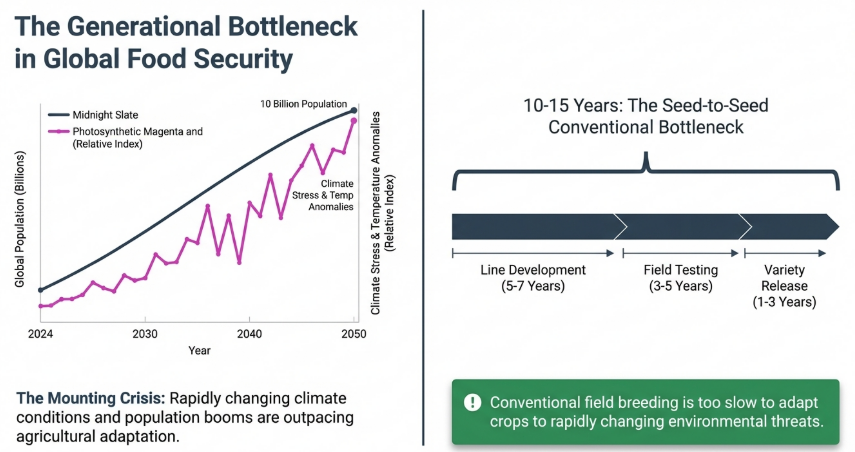
The global agricultural sector faces an existential “perfect storm.” With the world population projected to reach 10 billion by 2050, the demand for nutritional security is escalating against a backdrop of climate volatility and shifting pest dynamics. Traditionally, the primary bottleneck has been the 10–15 year duration of the breeding cycle. From a strategic management perspective, this sluggish pace represents a massive R&D overhead and a delayed Return on Investment (ROI) that modern food systems can no longer sustain. Speed Breeding (SB) serves as a critical savior, offering a technological leap to bypass biological constraints.
Originating from NASA-inspired research to grow wheat (variety ‘USU-Apogee’) in space stations, the technique was formalized for terrestrial application by researchers at the University of Queensland in 2003. SB is a suite of environmental manipulation techniques—often termed Phyto-Engineering—that “hacks” plant physiology to accelerate flowering and seed set. By transitioning from unpredictable open-field environments to Controlled Environment Ecosystems (CEE), SB allows for the rapid advancement of generations as quickly as modified physiology permits.
The Mechanism: What is Speed Breeding?
Speed Breeding (SB) is not a singular tool but a suite of techniques under the umbrella of Rapid Generation Advancement (RGA). It involves the meticulous manipulation of environmental conditions to accelerate flowering and seed set, allowing researchers to reach the next generation as quickly as possible.
Core Principles of Speed Breeding:
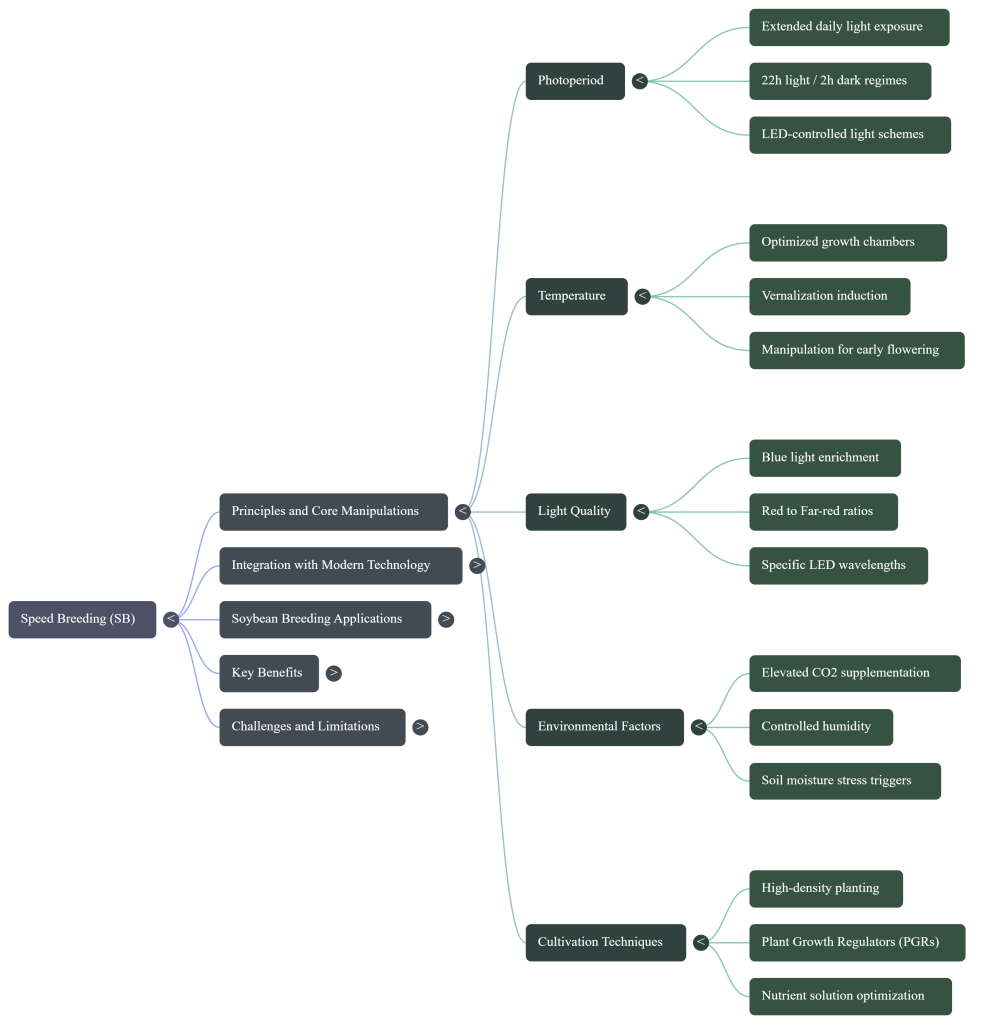
- Environmental Manipulation: Adjusting photoperiod, temperature, and CO2 to “trick” the plant into rapid development.
- Single Seed Descent (SSD): This is the preferred method for SB because it maximizes genetic diversity while minimizing the physical space required for each generation, as only one seed per plant is needed to advance.
- Precision Control: Using growth chambers to maintain stable artificial ecosystems 24/7.
While techniques like Double Haploid (DH) technology can create homozygous lines quickly, Speed Breeding has apexed them all. DH technology is highly genotype-dependent and requires a high level of technical skill. In contrast, SB is far more flexible and broadly applicable across diverse varieties once the protocol is optimized.
Moving from the plant’s “eyes” (light) to its “metabolic engine” (temperature), we find the specific levers used to control these biological clocks.
2. Comparative Analysis: Traditional Breeding vs. Speed Breeding
The fundamental barrier to innovation is the “seed-to-seed” cycle. Traditional breeding remains tethered to natural seasonality, creating a rigid innovation ceiling. Speed Breeding deconstructs this limitation, creating a “perpetual summer” that drastically alters the competitive landscape for seed developers.

| Aspect | Traditional Breeding | Transgenic Breeding | Speed Breeding |
|---|---|---|---|
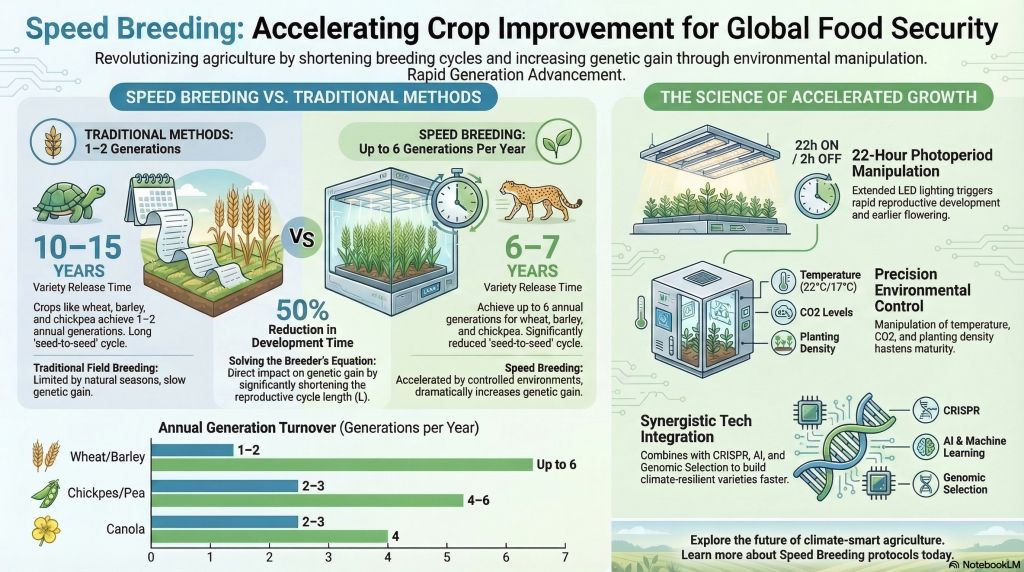
| Cycle Duration | 10–15 years | 4–5 years | 2–3 years |
| Methodology | Natural pollination/seasons | Gene manipulation/Open field | Controlled Environment (Light, Temp, CO2) |
| Resource Needs | Land-intensive (High acreage) | Medium land requirements | Infrastructure-intensive (CEE) |
| Genetic Gain | Gradual/Incremental | Targeted but slow testing | Accelerated/Rapid gain per year |
| Evaluation | 1–2 generations annually | 1–2 generations annually | Up to 10 generations annually |
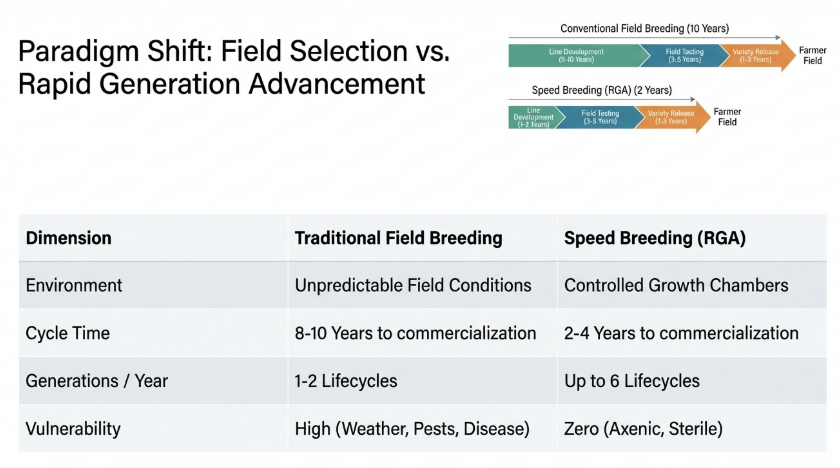
Strategic Interpretation: The reduction of line development from 10 years to 2 years enables “agile trait introgression.” For research directors, this means the ability to respond to emerging biosecurity threats in real-time, ensuring that a variety’s genetics remain relevant to field pressures at the moment of commercial release.
3. The Core Mechanisms of Environmental Manipulation
Plant development is a plastic response to external cues. SB manipulates these signals to maximize vegetative and reproductive transitions.
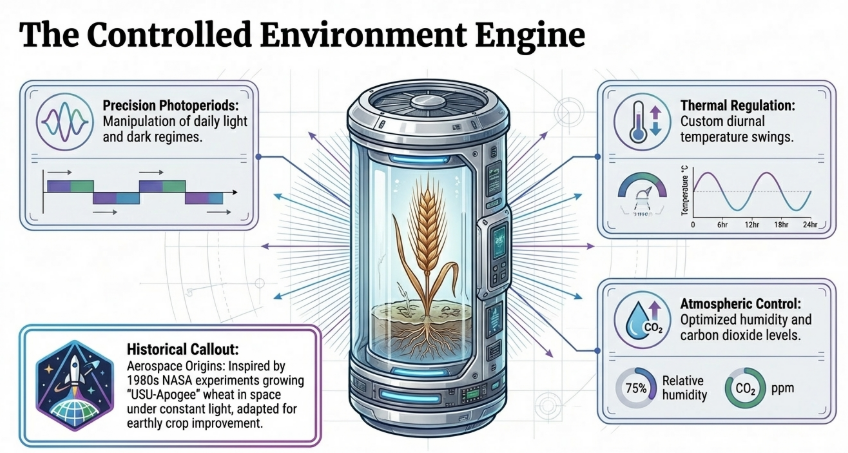
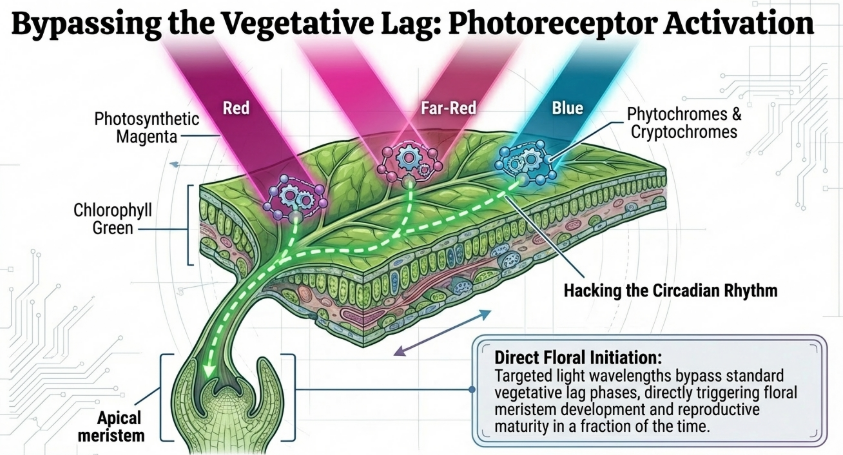
3.1 Photoperiod (Lighting the Path)
The photoperiod—the daily exposure to light and dark—is a sophisticated signal that tells the plant’s internal clock when to transition from growth to reproduction. Light is not just for photosynthesis; it is a developmental trigger.
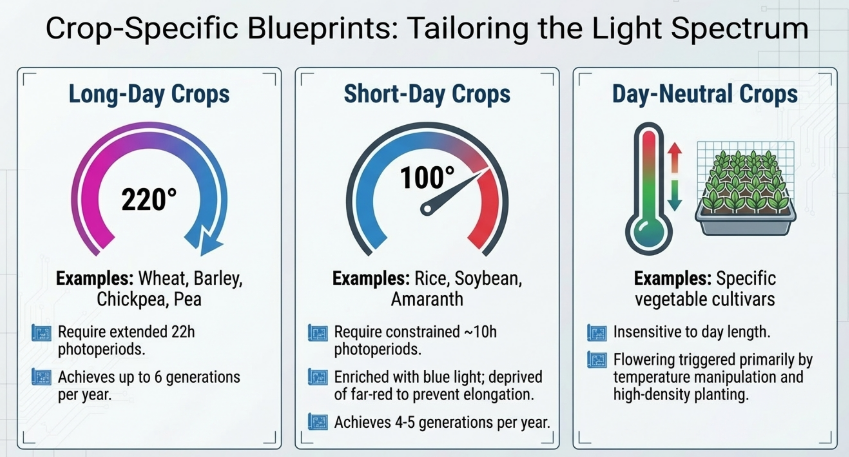
- Long-Day Crops (e.g., Wheat, Barley): These crops flower faster when exposed to extended days.
- Short-Day Crops (e.g., Soybeans): These crops require long nights to flower. To speed them up, scientists use specialized LED setups.
Research Insight: The Wheat and Soybean Contrast For wheat, a study by Dubcovsky et al. (2006) observed that a photoperiod of 22 hours light and 2 hours dark reduced the total number of days to flowering by half compared to standard 12/12-hour cycles. Conversely, for short-day crops like soybeans, researchers utilize a 10-hour photoperiod with blue-light enriched and far-red deprived LED schemes to achieve sturdy plants that mature in just 77 days.
By mastering light, we maximize energy while constantly signaling the plant that it is “time to reproduce.”

3.2 Temperature (The Thermal Catalyst)
Temperature acts as the catalyst for the plant’s internal chemical reactions. In a controlled environment, we provide stable thermal regimes that nature cannot guarantee, utilizing day/night cycles such as 22/17°C or 25/22°C.
💡 Pro-Tip: Vernalization While we generally provide optimal growth temperatures, breeders use strategic cooling—known as vernalization—to mimic winter. This cold treatment is essential for certain crops to trigger the transition from the vegetative stage to the reproductive flowering stage.
These stable environments protect plants from the unpredictable heatwaves or cold snaps that characterize open-field farming. Moving from the thermal engine to the atmospheric fuel, we look at CO2 levels.
3.3 CO2 Levels (The Atmospheric Boost)
Elevating Carbon Dioxide (CO2) levels in a sealed chamber to 400–700 ppm puts the plant’s growth on “turbo” by facilitating a faster shift to the reproductive stage.
Significant Impacts of CO2 Supplementation:
- Reduction in Flowering Time: In soybeans, elevated CO2 can reduce the days to flowering by approximately two days.
- Increased Seed Yield: Supplemental CO2 significantly increases the average seed number and weight per plant.
- Enhanced Crossing Efficiency: CO2 supplementation produces a higher count of healthy flowers, providing more opportunities for manual pollination.
The “So What?” for the Breeder: Beyond faster growth, CO2 supplementation allows for the miniaturization of the breeding setup. Studies (e.g., Nagatoshi & Fujita) show that healthy flowers can be produced even in smaller 0.4L pots. This means breeders can fit higher plant populations into smaller growth chambers, drastically increasing the scale of the breeding program.
As we automate these levers, we turn to the modern technological toolkit.
3.4 Soil Moisture Stress: Controlled drought stress is utilized strategically to trigger early maturation. In cowpea, this manipulation can induce flowering up to 12 days earlier than well-irrigated controls.
3.5 High-Density Planting (HDP) Dynamics In SB, HDP (up to 400 plants m−2) induces intense resource competition. This triggers a “shade avoidance” response, fast-tracking plants into reproductive stages. When combined with managed moisture stress post-flowering, the seed-to-seed cycle is compressed while maintaining seed viability. This physical infrastructure must be maintained with high-precision stability to sustain such biological acceleration.
3.6 Nutrient and Hormone Delivery: The exogenous application of Plant Growth Regulators (PGRs) such as Gibberellins and Cytokinins is used to break seed dormancy and expedite seed set, ensuring the high metabolic demands of the accelerated cycle are sustained.
“In 2017, the University of Queensland, Australia produced the first speed-bred spring wheat variety, named ‘DS Faraday’, showcasing the power of these techniques to deliver disease-resistant varieties in record time.”
4. Case Studies in Crop-Specific Acceleration
SB requires “crop-specific optimization” to account for the unique physiological triggers of different species. Recent synthesis of global data (Sharma et al. 2024) reveals the diverse potential of the platform:
- Rice (Oryza sativa): Using the “SpeedFlower” protocol (14h light/10h dark initial cycles), breeders achieve 4–5 generations per year.
- Soybean, Pea, and Oat: Achieve 5 generations per year. For soybeans, a 10-hour photoperiod enriched with blue LEDs and deprived of far-red light prevents excessive petiole elongation and lodging.
- Cereals (Wheat and Barley): Optimized 22-hour light regimes consistently deliver 6 generations per year.
- Pulses (Chickpea and Lentil): Chickpeas reach 6–7 generations, while lentils can achieve up to 8 generations annually through intensive light manipulation.
- Pigeonpea: A traditionally long-duration crop has been accelerated using a tailored photoperiod strategy of 13h:8h:13h at the vegetative, flowering, and pod-filling stages, respectively.
- Arabidopsis: The model plant can reach an unprecedented 10 generations per year.
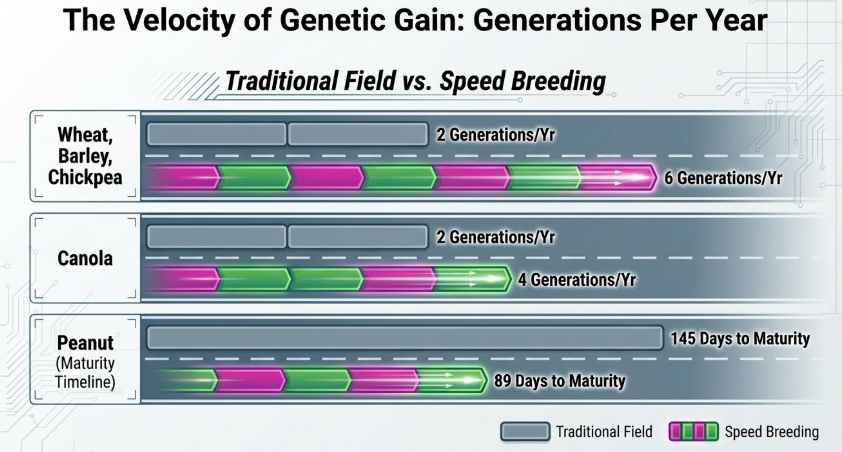
Generations Per Year: A Comparison
| Crop Type | Traditional (Field) | Speed Breeding (CEE) |
|---|---|---|
| Soybean | 1–2 generations | 5 generations |
| Wheat | 1–2 generations | 6–7 generations |
| Pea | 2–3 generations | 6 generations |
| Chickpea | 2–3 generations | 4–7 generations |
| Lentil | 1 generation | 8 generations |
| Arabidopsis | 1–2 generations | 10 generations |
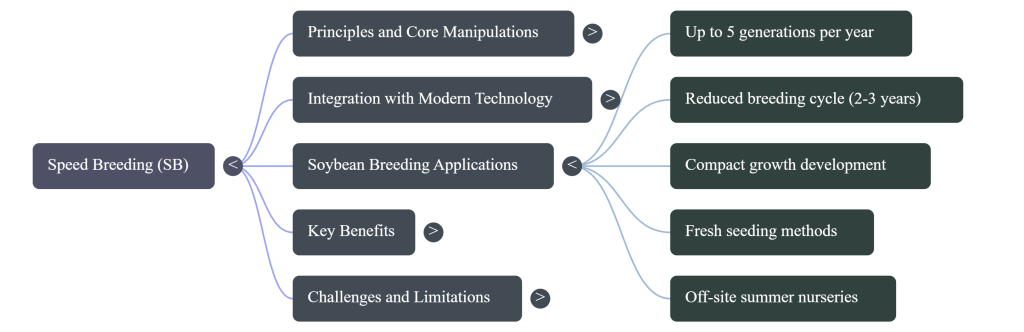
Case Study: Accelerated Breeding in Soybean (Glycine max)
As the world’s primary oilseed, accounting for over 60% of global production, soybean is vital for food and industrial security. However, soybean breeding is traditionally hindered by “genotype inflexibility” and a low transformation frequency (typically <5%). SB protocols specifically address these bottlenecks to compress the breeding window.
The Soybean SB Protocol
The protocol utilizes a blue-light-enriched and far-red-deprived LED scheme with a 10h photoperiod. This specific spectrum is critical: blue light leads to short, sturdy plants, while the deprivation of far-red light prevents lodging and excessive petiole elongation. Under this regime, soybean can flower as early as 23 Days After Sowing (DAS) and reach maturity within 77 days. To further shorten the cycle, the “Fresh Seeding Method” is employed, where mature pods are harvested at the R6 (full-seed) stage, artificially dried, and immediately sown, saving 7–10 days per generation and circumventing genotype-dependent maturation delays.
Soybean Application Outcomes
| Cultivation Condition | Focus/Parameter | Key Results |
|---|---|---|
| Blue-enriched/FR-deprived LED | Growth & Maturity | Flowering @ 23 DAS; maturation within 77 days; 5 generations/year. |
| Red and Blue (RB) Light | Morphology | Shortened planting-to-R7 interval by 1.5 days; significant height reduction vs. Full Spectrum (FS). |
| CO2 (400–700 ppm) | Crossing Efficiency | Significantly higher flower count; 98.5% hybrid success rate (Cultivar Enrei). |
| Fresh Seeding (R6 Stage) | Generation Turnover | Circumvents genotype flexibility issues; shortens cycle by 7–10 days. |
This integrated methodology allows for 5 generations per year, a 500% increase in throughput compared to traditional field systems.
Precision Protocols for Glycine max
Soybeans are uniquely sensitive to environmental cues, particularly photoperiod and temperature. SB triggers the rapid transition from vegetative growth to reproductive maturity by optimizing external signals, thereby shortening the interval to flowering and seed set.
Light Spectrum and Photoperiod Architecture Optimization requires a balance of light quality and duration to prevent excessive vegetative delay. A lower Red:Blue (RB) ratio is critical; research indicates this reduces plant height at the R1 and R7 stages and shortens the interval to maturity by 1.5 days compared to Full Spectrum (FS) lighting.
| Parameter | Specifications | Biological Impact |
|---|---|---|
| Photoperiod | 10-hour light / 14-hour dark | Triggers rapid flowering response in short-day genotypes. |
| Spectral Ratio | Low Red:Blue (RB) ratio | Shortens planting-to-maturity (R7) by 1.5 days; produces sturdy plants. |
| Light Quality | Blue-light enriched; Far-red deprived | Prevents petiole elongation and lodging; far-red light negatively impacts morphology. |
| Light Intensity | >1000μmol/m2s | Accelerates flowering by ~2 days; reduces overall plant stature for high-density setups. |
Atmospheric and Thermal Control Regimes
- CO2 Enrichment: Elevating CO2 to 400 ppm alone can reduce days to flowering by approximately two days. Supplementation up to 700 ppm further increases total flower count and crossing efficiency, enabling higher hybrid volume.
- Thermal Setpoints: A regime of 25/22∘C (Day/Night) is maintained. These setpoints are strategically selected to ensure meiotic progression stability, preventing the disruptions and embryo abortions common in thermal extremes.
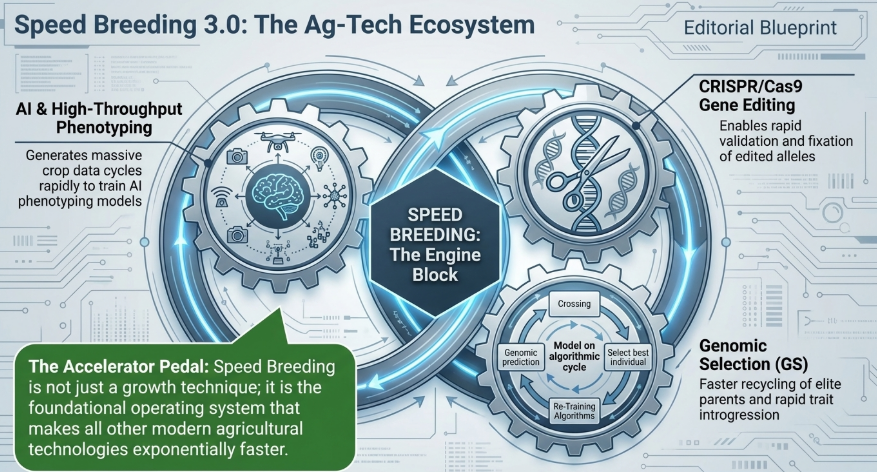
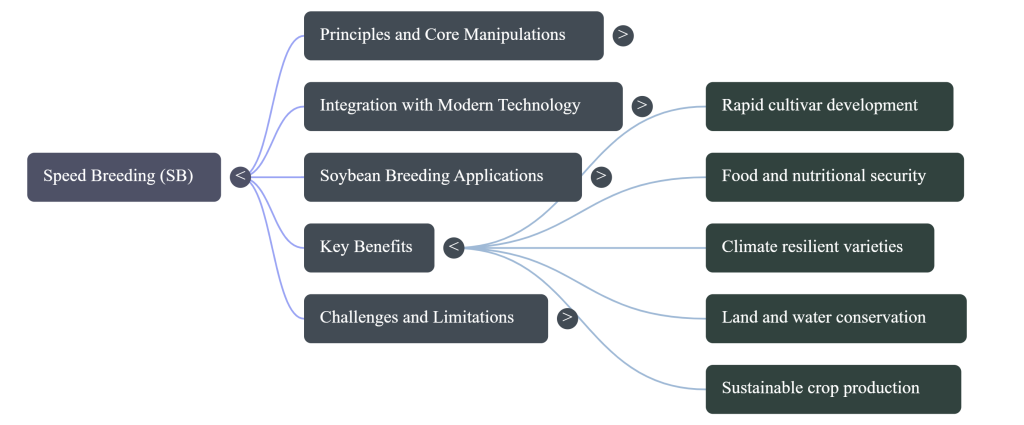
5. The “Speed Breeding Plus” Layer: Synergistic Biotech Integration
SB is a force-multiplier when integrated with molecular and digital tools, maximizing the “Genetic Gain” per unit of time.
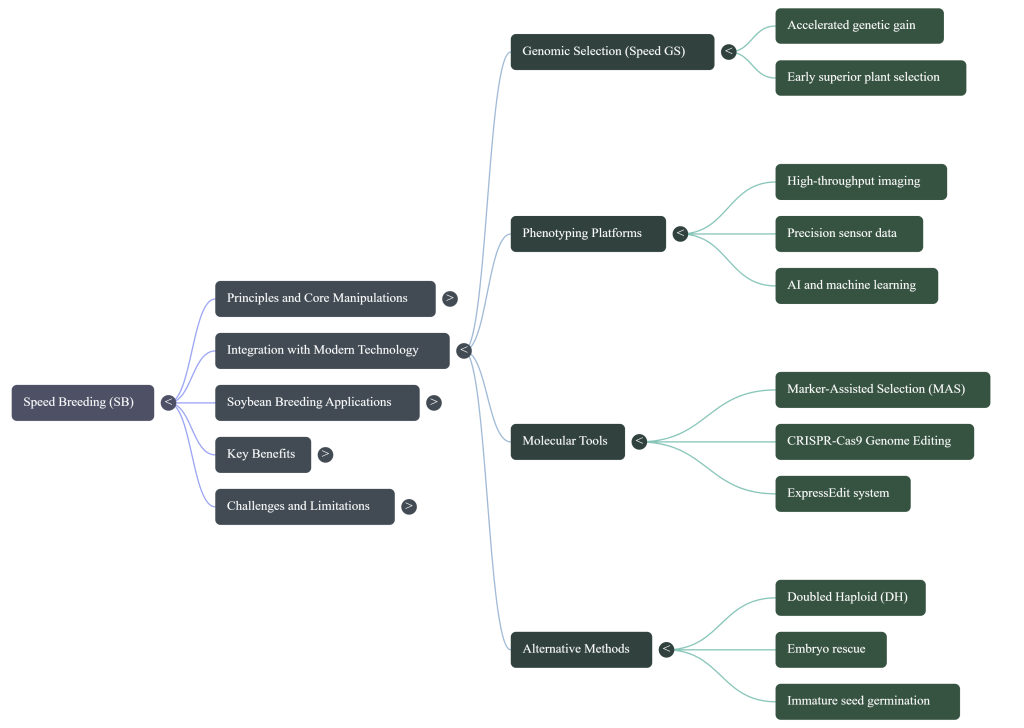
- Speed Genomic Selection (Speed GS): Superior plants are identified at the seedling stage within an accelerated cycle, allowing only elite genetics to advance.
- ExpressEdit: This is a pinnacle of integration, combining Marker-Assisted Selection (MAS) and preassembled CRISPR tools. Crucially, it bypasses the tissue culture and callus induction stages, which are traditionally major time-sinks and sources of genotype-dependency.
- AI & High-Throughput Phenotyping (HTP): Modern CEEs use Neural Networks and Mechanistic and Physiological Models (MPM-ML) to monitor and regulate Nutrient Use Efficiency (NUE) and Water Use Efficiency (WUE) through sensor-based data, replacing subjective manual observation.
Architecture of Controlled Environment Ecosystems (CEE)
CEE (or Controlled Environment Agriculture) represents the sophisticated infrastructure required to host SB. These chambers provide total climate stability, offsetting greenhouse gas emissions and delivering accurate nutrient ions more precisely than the unpredictability of traditional soil-based agriculture.
CEE includes diverse soilless technologies: Hydroponics, Aeroponics, and Aquaponics. These systems are managed through advanced computational intelligence models to maintain metabolic efficiency.
Computational Intelligence Models in CEE
| Model | Application & Impact |
|---|---|
| MPM-ML (Mechanistic/Physiological with Machine Learning) | Evaluates nutrient/water dynamics, photosynthetic assimilation products, and carbon sequestration. |
| RNN-LSTM (Recurrent Neural Network) | Deep learning model for the continuous monitoring of pH, temperature, and humidity. |
| Barber-Cushman Model | Calculates precise nutrient concentrations and mass transfer in solutions to optimize ion delivery. |
CEE architectures provide a stable artificial ecosystem that conserves 90% of water and 80% of land usage compared to traditional agriculture, while providing the precise control necessary to prevent yield loss from climatic instability.
Comparison of Selection Methods in SB
| Method | Application in SB | Maintenance of Diversity |
|---|---|---|
| Single Seed Descent (SSD) | Primary RGA method; rapidly achieves homozygosity. | High; preserves all alleles until fixation. |
| Single Pod Descent (SPD) | Condensed maintenance of multiple seeds per line. | Moderate. |
| Single Plant Selection (SPS) | Targeted selection in segregating generations. | Lower; risks allele loss without molecular markers. |
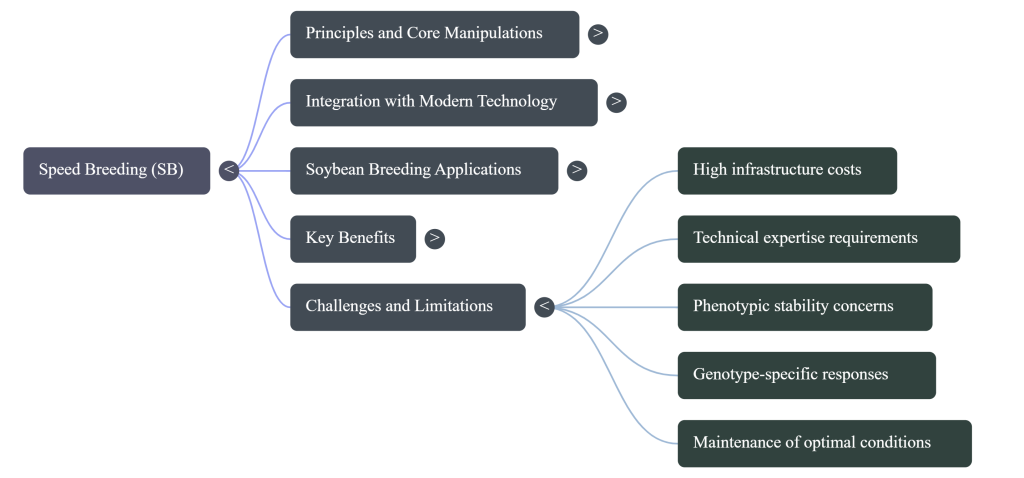
6. Implementation Challenges and Economic Realities
While transformative, the entry barriers to SB remain significant for resource-constrained programs.
- Biological Risks: Continuous light exposure can lead to photo-oxidation, chlorosis, and leaf injury. Phenotypic expression in a chamber may also vary from the field, requiring protocol refinement to ensure trait stability. A primary risk is the “cross-over interaction.”
- Genotype Sensitivity: Differential responses across cultivars mean that protocol optimization must be crop-specific; a protocol for bread wheat (Triticum aestivum) cannot be universally applied to soybean (Glycine max).
- Infrastructure and Energy: High initial costs for LED arrays and growth chambers are exacerbated by constant electricity demands. To democratize this technology, global research consortiums are now advocating for solar-powered or battery-powered inverter systems to sustain SB operations in developing nations.
- Expertise Gaps: The requirement for specialized technical skills in bioinformatics and environmental engineering necessitates international collaboration to standardize protocols across diverse crop species.

Failure Mitigation Roadmap (Standard Operating Procedure)
- Biosecurity: Maintain axenic/sterile environments to prevent pest emergence; utilize HES-driven sensors for early stress detection.
- System Redundancy: Deploy solar/battery backups for air conditioning to prevent heat-stress-induced embryo abortion during power fluctuations.
- Seed Viability Protocol: Harvest seeds at the R6 (full seed) stage, followed by artificial drying. This prevents dormancy issues and allows for “immediate re-sowing,” the linchpin of achieving 5+ generations per year.
Strategic Barriers & Mitigation Strategies
| Barrier | Institutional Risk | Mitigation Strategy |
|---|---|---|
| High Capital Expenditure (CAPEX) | Cost of LEDs and intensive climate control. | Solar integration and inverter-based cooling systems to reduce energy footprint. |
| Phenotypic Stability | Traits expressed in CEE may not correlate with the field. | Routine field validation and refined HTP protocols for cross-environment correlation. |
| Technical Expertise | Multidisciplinary skill gap (bioinformatics/engineering). | Collaborative research networks and intensive cross-training programs. |
| Biological Constraints | Seed viability in rapid cycles. | Fresh Seeding: Harvesting at R6 (full-seed) stage followed by immediate sowing; embryo rescue. |
7. Strategic Directive: The Next Breakthrough
Speed Breeding is a paradigm shift that reduces genetic progression time by 50–70%. It is no longer an optional innovation but a strategic imperative for global research institutions tasked with achieving “Zero Hunger.” To fulfill the promise of a sustainable 10-billion-person food system, it is essential that we standardize these protocols, integrate them with ExpressEdit platforms, and utilize renewable energy to ensure this technology is accessible worldwide.
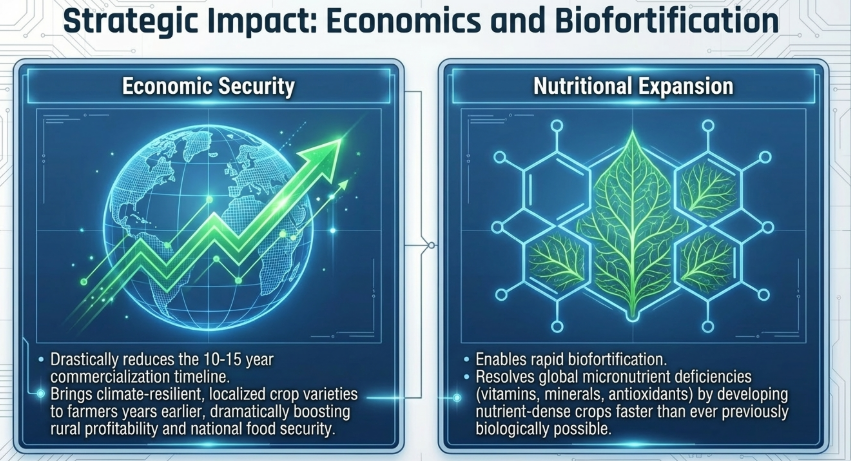
Image Summary
Questions/Answers
1. How does speed breeding reduce long traditional crop development cycles?
Speed breeding reduces long traditional crop development cycles by manipulating the growing environment to accelerate a plant’s natural life cycle, enabling researchers to achieve multiple generations in a single year. While conventional breeding typically takes 10 to 15 years to develop and release a new variety, speed breeding can condense this timeline to 6 to 7 years by drastically shortening the time required for line development and fixation.
This reduction is achieved through several core environmental and physiological interventions:
1. Manipulation of Light and Photoperiod
The primary driver of speed breeding is the use of extended photoperiods, often providing plants with up to 22 hours of light followed by a brief 2-hour dark period. This near-continuous exposure to light accelerates photosynthesis and triggers early floral initiation, particularly in long-day plants like wheat, barley, and peas. Modern protocols also utilize LED lighting to optimize light quality, such as blue and far-red spectrums, which can further hasten the transition from vegetative to reproductive stages.
2. Precise Temperature and Humidity Control
Speed breeding facilities maintain optimal temperature regimes tailored to specific crops to accumulate Growing Degree Days (GDD) more rapidly. Elevating temperatures within a crop’s safe physiological limits facilitates faster development and maturity. Additionally, speed vernalization techniques integrate cold treatments into the accelerated growth cycle, allowing winter-type crops that normally require months of winter chilling to flower and produce seeds much faster.
3. Early Seed Harvest and Maturation
Traditional breeding requires seeds to mature naturally on the plant, a process that can take weeks. Speed breeding shortens this by harvesting immature seeds (often just 14–20 days after flowering) and ripening them artificially in specialized dehydrators or ovens. This “seed-to-seed” shortcut can save several weeks per generation. In some cases, embryo rescue is used, where young embryos are extracted and grown in vitro to bypass seed dormancy entirely.
4. Advanced Management Strategies
- High-Density Planting: Growing plants in close proximity creates competition for light and nutrients, which induces stress and can trigger earlier flowering as an escape mechanism.
- Single Seed Descent (SSD): Speed breeding often employs SSD, where only one seed is advanced per plant, allowing for the rapid generation of homozygous lines (genetically stable plants) in compact, resource-efficient growth chambers.
- CO2 Enrichment: Supplementing growth environments with elevated carbon dioxide levels (e.g., 400–800 ppm) can further enhance biomass accumulation and speed up the transition to the reproductive phase in certain crops like soybeans and rice.
5. Integration with Modern Technology
The impact of speed breeding is amplified when integrated with other biotechnological tools:
- Genomics and Marker-Assisted Selection (MAS): Breeders can use DNA markers to select for desirable traits as early as the seedling stage, ensuring that only the best plants are advanced through the rapid cycles.
- Genome Editing (CRISPR/Cas9): Combining speed breeding with gene editing allows for the rapid fixation and evaluation of new genetic modifications across multiple generations in a single year.
- Artificial Intelligence (AI): AI is increasingly used to analyze the massive amounts of data generated by speed breeding programs and to optimize growth parameters for even faster results.
2. Explain the role of CO2 supplementation in speed breeding.
In speed breeding, carbon dioxide (CO2) supplementation acts as a physiological lever to accelerate photosynthesis and promote rapid plant development, thereby shortening the overall breeding cycle. While not considered a fundamental requirement like light and temperature manipulation, it is a potent supplementary tool used to enhance growth rates and reproductive efficiency.
The role of CO2 supplementation in speed breeding includes:
1. Enhancement of Photosynthesis and Biomass
CO2 is the primary substrate for photosynthesis. By elevating CO2 levels—typically from the ambient ~400 ppm to between 800 and 1500 ppm—researchers can significantly increase leaf photosynthetic rates. This leads to:
- Increased Biomass: Enhanced total leaf area, plant height, and dry weight of stems, roots, and leaves.
- Stronger Growth: CO2 enrichment provides a steady supply of assimilates that sustain the accelerated growth required by speed breeding’s intensive lighting and temperature regimes.
2. Acceleration of Reproductive Transitions
Elevated CO2 concentrations can hasten the transition from the vegetative to the reproductive stage, leading to earlier flowering and heading.
- Soybean: In combination with optimized light and temperature, CO2 supplementation has been shown to reduce the crop cycle of certain cultivars from 102–132 days down to just 70 days.
- Rice: Increased CO2 levels (e.g., 600 ppm) have reduced the days to heading by approximately 3 to 9 days, depending on the cultivar.
- Cowpea: Some studies reported flowering occurring up to 12 days earlier under enriched CO2.
3. Improved Crossing and Yield Components
Beyond just speed, CO2 supplementation improves the quality of the breeding material:
- Flower Production: It consistently increases the number and size of healthy flowers, which enhances crossing efficiency for breeders.
- Seed Quality: Supplementation can lead to a higher number of seeds per plant and increased average seed weight, ensuring robust genetic material for the next generation.
4. Species-Specific Variations (C3 vs. C4)
The effectiveness of CO2 supplementation is highly dependent on the plant’s photosynthetic pathway and genotype:
- C3 Plants: Crops like wheat, rice, and soybean are more sensitive to CO2 enrichment and typically show a more pronounced growth response.
- C4 Plants: Crops such as maize and pearl millet show a less significant response because their internal CO2-concentrating mechanisms are already near saturation at ambient levels. In some C4 species like pearl millet, CO2 may increase biomass without accelerating flowering, and in some cases, it may even delay it.
5. Infrastructure and Safety
Implementing CO2 supplementation requires fully enclosed growth chambers or retrofitted glasshouses equipped with CO2 cylinders, regulators, and analyzers. This adds to the operational and capital costs of a breeding facility. Additionally, practitioners must strictly adhere to health and safety protocols when handling CO2 gas systems.
3. What environmental factors are manipulated to trigger rapid plant growth?
Speed breeding triggers rapid plant growth and development by meticulously manipulating several core environmental factors within controlled-environment ecosystems. By optimizing these variables, breeders can bypass seasonal constraints and achieve multiple generations in a single year.
The primary environmental factors manipulated include:
1. Light Management (Photoperiod, Quality, and Intensity)
Light is the most critical factor in speed breeding, acting as both an energy source and a developmental signal.
- Extended Photoperiod: Most speed breeding protocols use a 22-hour light period followed by a 2-hour dark phase to support the circadian clock while maximizing photosynthesis. For long-day plants, this triggers early flowering by simulating continuous summer conditions.
- Light Quality (Spectrum): Breeders use specialized LED lighting to focus on the blue, red, and far-red wavelengths (400–700 nm). Blue light influences stomatal behavior and compact growth, red light activates phytochromes to regulate flowering, and far-red light can induce “shade avoidance” responses that accelerate floral transitions.
- Light Intensity: Standard protocols recommend a Photosynthetic Photon Flux Density (PPFD) of 360–650 µmol/m²/s, which provides sufficient energy for rapid biomass accumulation.
2. Temperature and Humidity Regulation
Precision control over the thermal and moisture environment prevents stress and accelerates metabolic rates.
- Temperature Regimes: A common regime is 22°C during the day and 17°C at night. Higher temperatures can be used to speed up vegetative growth, while cooler night temperatures help plants recover from the stress of near-continuous light.
- Vernalization: For winter crops that require chilling to flower, “speed vernalization” involves exposing germinated seeds to low temperatures (approx. 10°C) under extended light for 4–6 weeks to bypass natural winter cycles.
- Relative Humidity (RH): Humidity is typically maintained between 60% and 70% to prevent plant desiccation and ensure adequate transpiration while minimizing the risk of fungal diseases.
3. Atmospheric Composition (CO2 Supplementation)
Supplementing growth chambers with elevated carbon dioxide (CO2) levels—typically ranging from 400 to 1500 ppm—can significantly boost the rate of photosynthesis. This enrichment enhances biomass production and, in certain crops like rice and soybean, shortens the time required to reach the reproductive stage.
4. Plant Density and Competition
Growing plants at high densities (e.g., up to 1000 plants per square meter) induces competition for light and nutrients. This competition acts as a form of physiological stress that can trigger earlier flowering as a survival “escape mechanism,” allowing for rapid generation turnover in compact spaces.
5. Nutrition and Hormone Delivery
- Soil and Nutrients: Precise delivery of macro and micronutrients supports the rapid metabolic demands of accelerated growth. Reducing soil moisture or nutrients after flowering can also be used to hasten grain filling and maturation.
- Plant Growth Regulators (PGRs): Synthetic hormones like auxins, cytokinins, and gibberellins are sometimes applied to induce earlier flowering, improve seed set, or manage plant height in restricted spaces. For instance, gibberellic acid (GA3) can be used to break seed dormancy and promote uniform germination.
4. What are the economic challenges of maintaining speed breeding facilities?
The economic challenges of maintaining speed breeding facilities are significant, primarily centered around high initial setup costs, substantial energy demands, and the requirement for continuous, specialized resource management. These financial barriers are often the most acute for public sector programs and researchers in developing nations.
1. High Initial Capital Investment
Establishing a functional speed breeding facility requires a large upfront investment in infrastructure.
- Sophisticated Infrastructure: Facilities must be equipped with temperature-controlled glasshouses or fully enclosed growth chambers. Growth chambers alone can cost tens of thousands of dollars, which is a major barrier for many research projects.
- Facility Setup Costs: The capital investment for a controlled environment plant growing chamber (approx. 500–1000 square feet) can range from 20 to 30 lakh Indian rupees.
- Specialized Equipment: Beyond the structure, costs include carbon dioxide analyzers, relative humidity controllers, high-end sensors, and specialized lighting systems.
2. Substantial Operational and Energy Costs
The ongoing expense of running these facilities is a primary economic hurdle, often exceeding the establishment costs over time.
- Energy Consumption: Maintaining the intensive 22-hour photoperiods and precise temperature regimes (heating in winter, cooling in summer) is highly energy-intensive.
- Major Cost Driver: Lighting and temperature control typically account for more than 50% to 60% of the total operational costs. For example, temperature regulation alone accounted for over half of plant management costs in certain Australian winter trials.
- Resource Inputs: Facilities require a continuous and reliable supply of electricity, distilled water, and nutrient solutions, all of which add to recurring expenses.
3. Human Capital and Personnel Expenses
Maintaining these systems requires a high level of technical expertise.
- Skilled Labor: Speed breeding demands trained plant breeders and technicians who understand agronomy, crop physiology, and the operation of complex climate-control technology.
- Staff Retention: Public sector programs face a “brain drain,” where highly trained personnel frequently migrate to private seed companies that offer better remuneration.
4. Regional and Institutional Constraints
Developing nations and public programs face systemic economic challenges.
- Infrastructure Gaps: Many regions lack the reliable electricity and water grids necessary for the “24/7” nature of speed breeding.
- Funding Instability: These programs require sustained, long-term financial support for R&D, which is often missing in the public sector where institutional support is limited.
5. Economic Recoupment and ROI
Despite high costs, speed breeding can be a cost-effective strategy in the long term.
- Time-to-Market: By reducing variety development time from 10–15 years to 6–7 years, organizations can release new cultivars faster.
- Commercialization: Linking speed breeding programs to systems for rapid commercialization allows organizations to recoup the extra breeding expenses more quickly than conventional methods.
- Resource Efficiency: Because plants are grown at higher densities in smaller spaces, speed breeding can optimize land and labor use.
5. How does speed breeding compare to doubled haploid technology?
Speed breeding (SB) and doubled haploid (DH) technology are both advanced methods used to shorten the traditionally long breeding cycles of crops, but they achieve this through different biological and technical means. While DH technology achieves complete homozygosity (genetic stability) in just one or two generations, speed breeding uses environmental manipulation to accelerate the natural life cycle, achieving multiple generations per year.
Key Differences and Comparisons
| Feature | Speed Breeding (SB) | Doubled Haploid (DH) |
|---|---|---|
| Method | Environmental manipulation (light, temp, humidity). | Haploid embryo rescue and chromosome doubling. |
| Generations/Year | Typically 4–6 (up to 9 in some crops). | Reaches homozygosity in 1–2 generations. |
| Time to Pure Line | ~1 to 2 years for most cereals and legumes. | ~1 to 1.5 years for most genotypes. |
| Recombination | Allows multiple rounds of meiosis/recombination. | Restricts recombination to a single round of meiosis. |
| Genotype Sensitivity | Highly versatile; applicable to diverse germplasm. | Highly genotype-dependent; variable success rates. |
| Infrastructure | Enclosed growth chambers/glasshouses. | Specialized tissue culture labs and equipment. |
Advantages of Speed Breeding over DH
- Recombination Frequency: Because SB involves multiple generations of self-pollination, it allows for more meiotic events. This results in a higher recombination frequency, which is a major advantage for genetic mapping and breaking undesirable gene linkages.
- Versatility: SB is suitable for diverse germplasm without the need for specialized laboratory conditions for in vitro tissue culture.
- Plant Vigor and Survival: SB avoids common DH hurdles such as low induction rates, poor plant vigor, low germination rates, and abnormal development (e.g., albinism) that affect some species.
- Cost-Effectiveness: In some species, SB can be a cheaper alternative to producing DH populations, especially when dealing with large segregating populations.
Advantages of DH over Speed Breeding
- Speed to Homozygosity: DH technology is generally considered the fastest method for reaching 100% homozygosity, as it bypasses the need for several generations of self-pollination.
- Uniformity: DH lines are 100% genetically uniform from the first generation, whereas SB lines typically reach approximately 98.4% homozygosity after six generations of selfing.
Synergistic Integration
The sources emphasize that these technologies are not mutually exclusive. Speed breeding can be used to further accelerate DH protocols by shortening the time required for initial parental crossing, plant regeneration, and the subsequent seed multiplication steps. When integrated with other tools like Marker-Assisted Selection (MAS) and Genomic Selection (GS), both methods significantly enhance genetic gain per unit of time.
6. How is speed breeding integrated with CRISPR gene editing?
The integration of speed breeding (SB) with CRISPR gene editing creates a powerful synergy that accelerates the development, fixation, and evaluation of edited traits. While CRISPR allows for precise genetic modifications, speed breeding provides the rapid generation turnover necessary to move these modifications from the laboratory into stable, breeding-ready lines in a fraction of the time required by traditional methods.
1. Key Integration Mechanism: ExpressEdit
A major advancement in this field is an approach known as “ExpressEdit.” Traditionally, gene editing requires labor-intensive and time-consuming in vitro tissue culture and plant regeneration. ExpressEdit circumvents these bottlenecks by:
- Direct Application: Applying CRISPR components (such as Cas9 and sgRNA sequences) directly to plant tissues, like shoot apical meristems, using techniques such as particle bombardment or biolistics.
- Rapid Advancement: The treated plants are immediately grown under speed breeding conditions. This allows for up to six generations per year, enabling researchers to quickly screen progeny for the desired trait.
- Non-Transgenic Selection: Through rapid cycling, breeders can select plants that possess the edited trait but have segregated away the Cas9 machinery, resulting in “transgene-free” plants that are genetically indistinguishable from those with natural mutations.
2. Benefits of the Integrated Approach
- Rapid Homozygosity: Edited plants grown under SB protocols reach reproductive maturity faster. This enables the achievement of genetic stability (homozygosity) for the edited allele within just 1–2 years, compared to the 8–10 years often required in conventional systems.
- Rigorous Quality Control: The controlled environment of SB facilitates high-throughput screening. Desirable lines can be preselected at the T1 generation, with a strict evaluation conducted at the T2 generation to ensure the elimination of off-target genotypes.
- Multi-Trait Pyramiding: CRISPR/Cas systems can target multiple sites simultaneously. When combined with SB, these multiple desired traits can be pyramided and fixed into a single elite background across several rapid generations.
3. Practical Applications in Major Crops
The sources document several successful integrations of speed breeding and CRISPR:
- Wheat: Researchers have edited genes associated with grain size, quantity, and drought resistance, using SB to rapidly advance these high-yield lines.
- Tomato: Integration has been used to reduce ethylene biosynthesis, which significantly extends the shelf life of the fruit. It has also been used to introgress genes like CAB-13 to improve tolerance to the continuous light used in SB facilities.
- Rice: CRISPR has been used to knockout genes responsible for late flowering, enabling these varieties to complete their life cycles even faster under SB protocols.
- Perennial Fruit (Apple): CRISPR has been used to edit the TFL1 (Terminal Flower 1) gene to shorten the long juvenile phase, with SB facilitating the rapid evaluation of these early-flowering trees.
4. Strategic Optimization
Beyond just fixing traits, CRISPR is used to optimize plants specifically for speed breeding facilities. For example, gene editing can be used to tune a plant’s circadian clock or sensitivity to photoperiod, making it more productive under the intensive 22-hour light regimes typical of speed breeding.
5. Challenges and Constraints
Despite its potential, the integration faces hurdles. The regulatory classification of CRISPR-edited crops varies significantly by country, and some regions (like the EU) classify them as GMOs, which can impede global adoption. Additionally, the fast-cycling conditions of SB could theoretically amplify unintended off-target effects or epigenetic changes, requiring multi-generational stability assessments to ensure the safety of the new varieties.
References
Aggarwal, G., Jeena, A. S., Mehra, K., Kumar, B., Kashyap, S., Yadav, D. K., Maurya, A. K., Venkatesh, S. C., Singla, P., & Bohra, A. (2025). Speed-bred crops for food security and sustainable agriculture. In Planta (Vol. 262, Number 2). Springer Science and Business Media Deutschland GmbH. https://doi.org/10.1007/s00425-025-04746-6
Ahtisham, M., & Obaid, Z. (2025). Enhancing Crop Improvement with Speed Breeding: Applications, Challenges, and Future Prospects. Premier Journal of Plant Biology. https://doi.org/10.70389/pjpb.100017
Bashir, A., Abbas, A., Li, X., Shi, Q., Niu, D., & Zhang, L. (2025). Harnessing light, photoperiod and temperature for accelerated flowering in speed breeding: Mechanisms, applications and crop diversity. In Journal of Plant Physiology (Vol. 311). Elsevier GmbH. https://doi.org/10.1016/j.jplph.2025.154548
Bhargava, K., Abhishek, E., Madhusudhan, B., Naveen, A., Akhil, V. S., Yadav, T. V., Mudhale, A., & Kumar, G. A. (2023). A Review of Rapid Generation Advancement (RGA) in Crop Improvement. International Journal of Plant & Soil Science, 35(7), 138–145. https://doi.org/10.9734/ijpss/2023/v35i72873
Blinkov, A. O., Kroupin, P. Y., Dmitrieva, A. R., Kocheshkova, A. A., Karlov, G. I., & Divashuk, M. G. (2025). Speed breeding: protocols, application and achievements. In Frontiers in Plant Science (Vol. 16). Frontiers Media SA. https://doi.org/10.3389/fpls.2025.1680955
Borgohain, T., Suma, R., Muttappagol, M., Saikia, B., Keithellakpam, A., Laskar, A., Hiremath, S. S., Basu, U., Velmurugan, N., Palakolanu, S. R., & Chikkaputtaiah, C. (2026). Precision breeding in a changing climate: unlocking resilience through omics and gene editing. In Functional & integrative genomics (Vol. 26, Number 1, p. 26). https://doi.org/10.1007/s10142-025-01796-7
Ćeran, M., Miladinović, D., Đorđević, V., Trkulja, D., Radanović, A., Glogovac, S., & Kondić-Špika, A. (2024). Genomics-assisted speed breeding for crop improvement: present and future. In Frontiers in Sustainable Food Systems (Vol. 8). Frontiers Media SA. https://doi.org/10.3389/fsufs.2024.1383302
Cha, J. K., Park, H., Choi, C., Kwon, Y., Lee, S. M., Oh, K. W., Ko, J. M., Kwon, S. W., & Lee, J. H. (2023). Acceleration of wheat breeding: enhancing efficiency and practical application of the speed breeding system. Plant Methods, 19(1). https://doi.org/10.1186/s13007-023-01083-1
Chaudhary, N., & Sandhu, R. (2024). A comprehensive review on speed breeding methods and applications. In Euphytica (Vol. 220, Number 3). Springer Science and Business Media B.V. https://doi.org/10.1007/s10681-024-03300-x
Chiurugwi, T., Kemp, S., Powell, W., & Hickey, L. T. (2019). Speed breeding orphan crops. In Theoretical and Applied Genetics (Vol. 132, Number 3, pp. 607–616). Springer Verlag. https://doi.org/10.1007/s00122-018-3202-7
Dasgupta, T. (2025). Speed Breeding: A Unique Technology for Accelerated Varietal Development. International Journal of Agriculture Environment and Biotechnology, 18(3). https://doi.org/10.30954/0974-1712.03.2025.7
Ghosh, S., Watson, A., Gonzalez-Navarro, O. E., Ramirez-Gonzalez, R. H., Yanes, L., Mendoza-Suárez, M., Simmonds, J., Wells, R., Rayner, T., Green, P., Hafeez, A., Hayta, S., Melton, R. E., Steed, A., Sarkar, A., Carter, J., Perkins, L., Lord, J., Tester, M., … Hickey, L. T. (2018). Speed breeding in growth chambers and glasshouses for crop breeding and model plant research. Nature Protocols, 13(12), 2944–2963. https://doi.org/10.1038/s41596-018-0072-z
Gudi, S., Kumar, P., Singh, S., Tanin, M. J., & Sharma, A. (2022). Strategies for accelerating genetic gains in crop plants: special focus on speed breeding. In Physiology and Molecular Biology of Plants (Vol. 28, Number 10, pp. 1921–1938). Springer. https://doi.org/10.1007/s12298-022-01247-8
He, R., Ju, J., Liu, K., Song, J., Zhang, S., Zhang, M., Hu, Y., Liu, X., Li, Y., & Liu, H. (2024). Technology of plant factory for vegetable crop speed breeding. In Frontiers in Plant Science (Vol. 15). Frontiers Media SA. https://doi.org/10.3389/fpls.2024.1414860
Jan, S. A., Tabassum, R., & Bashir, H. (2022). Speed breeding methods for soybean improvement: recent advances. Journal of Nutritional Health & Food Engineering, 12(2), 41–42. https://doi.org/10.15406/jnhfe.2022.12.00354
Kabade, P. G., Kumar, S., Kohli, A., Singh, U. M., Sinha, P., & Singh, V. K. (2025). Speed breeding 3.0: mainstreaming light-driven plant breeding for sustainable genetic gains. In Trends in Biotechnology (Vol. 43, Number 10, pp. 2462–2478). Elsevier Ltd. https://doi.org/10.1016/j.tibtech.2025.04.011
Katiyar, S. K., Das, R. R., Pazhamala, L. T., Bartholomé, J., Chandel, G., Bilaro, A., Asante, M. D., Iftekharuddaula, K. M., Islam, M. M., Yadaw, R. B., Verma, R., Srinivas, T., Yeshala, C. M., Abade, H., Raharinivo, V., & Musila, R. (2025). Accelerated breeding modernization: a global blueprint for driving genetic gains, climate resilience, and food security in rice. In Theoretical and Applied Genetics (Vol. 138, Number 12). Springer Science and Business Media Deutschland GmbH. https://doi.org/10.1007/s00122-025-05060-1
Kayess, M. O., Siddiqui, M. N., Gupta, D. R., Uddin, M. J., & Islam, T. (2025). Speed breeding enhances crop resilience and productivity in a changing climate. In Molecular Breeding (Vol. 45, Number 10). Springer Science and Business Media B.V. https://doi.org/10.1007/s11032-025-01588-8
Mir, J. I., Shafi, S., Verma, M. K., Raja, W. H., Nabi, S. U., Sharma, O. C., Shah, M. A., Prusty, R., Priya, J., Kuchay, M. A., Parveen, N., Lone, A. H., Yasmeen, S., Rashid, U., & Mansoor, S. (2026). Speed breeding in perennial fruit crops as a novel strategy to reduce generation period. In Physiology and Molecular Biology of Plants. Springer. https://doi.org/10.1007/s12298-026-01713-7
Mitache, M., Zeroual, A., Baidani, A., Bencharki, B., & Idrissi, O. (2026). A Comprehensive Review of Speed Breeding Based on Extended Photoperiod in Food Legumes. Journal of Plant Growth Regulation. https://doi.org/10.1007/s00344-025-12047-1
Pathania, R., Kumar, S., Chahal, A. S., Singh, A., Kaplex, A., & Sharma, A. (2025). Advancing agriculture through speed breeding: protocols, integration, achievements and challenges. In Euphytica (Vol. 221, Number 11). Springer Science and Business Media B.V. https://doi.org/10.1007/s10681-025-03630-4
Potts, J., Jangra, S., Michael, V. N., & Wu, X. (2023). Speed Breeding for Crop Improvement and Food Security. In Crops (Vol. 3, Number 4, pp. 276–291). Multidisciplinary Digital Publishing Institute (MDPI). https://doi.org/10.3390/crops3040025
Rai, K. K. (2022). Integrating speed breeding with artificial intelligence for developing climate-smart crops. Molecular Biology Reports, 49(12), 11385–11402. https://doi.org/10.1007/s11033-022-07769-4
Reddy Pogu, A. Shiny., & Soharu, A. (2025). Speed Breeding in Wheat Crop: A comprehensive Review. Journal of Advances in Biology & Biotechnology, 28(6), 235–246. https://doi.org/10.9734/jabb/2025/v28i62389
Revolutionizing Crop Production: The Imperative of Speed Breeding Technology in Modern Crop Improvement. (2024a). Crop Breeding, Genetics and Genomics, 6(2). https://doi.org/10.20900/cbgg20240003
Revolutionizing Crop Production: The Imperative of Speed Breeding Technology in Modern Crop Improvement. (2024b). Crop Breeding, Genetics and Genomics, 6(2). https://doi.org/10.20900/cbgg20240003
Rossi, N., Powell, W., Halliday, K., & Sharma, R. (2026). From photoperiod thresholds to photoperiod sensitivity: dual strategies for cost-effective speed breeding and climate-ready barley. Frontiers in Plant Science, 17. https://doi.org/10.3389/fpls.2026.1742787
Samantara, K., Bohra, A., Mohapatra, S. R., Prihatini, R., Asibe, F., Singh, L., Reyes, V. P., Tiwari, A., Maurya, A. K., Croser, J. S., Wani, S. H., Siddique, K. H. M., & Varshney, R. K. (2022). Breeding More Crops in Less Time: A Perspective on Speed Breeding. In Biology (Vol. 11, Number 2). MDPI. https://doi.org/10.3390/biology11020275
Selvamuthu, K., & Alagarasan, G. (2026). A process engineering framework for parameter space optimization in speed breeding. In Plant Communications. Cell Press. https://doi.org/10.1016/j.xplc.2026.101746
Sharma, A., Hazarika, M., Heisnam, P., Pandey, H., Devadas, V. A. S. N., Kesavan, A. K., Kumar, P., Singh, D., Vashishth, A., Jha, R., Misra, V., & Kumar, R. (2024). Controlled Environment Ecosystem: A Cutting-Edge Technology in Speed Breeding. In ACS Omega (Vol. 9, Number 27, pp. 29114–29138). American Chemical Society. https://doi.org/10.1021/acsomega.3c09060
Sharma, S., Kumar, A., Dhakte, P., Raturi, G., Vishwakarma, G., Barbadikar, K. M., Das, B. K., Shivaraj, S. M., Sonah, H., & Deshmukh, R. (2023). Speed Breeding Opportunities and Challenges for Crop Improvement. In Journal of Plant Growth Regulation (Vol. 42, Number 1, pp. 46–59). Springer. https://doi.org/10.1007/s00344-021-10551-8
Shreya, Kumar, V., & Arjoo. (2022). Speed Breeding : Accelerated Plant Breeding. Journal of Agriculture Research and Technology, Special(01), 36–39. https://doi.org/10.56228/jart.2022.sp107
Singh, S., & Nanda, S. R. (2024). Accelerating soybean breeding: A comprehensive review of speed breeding. International Journal of Advanced Biochemistry Research, 8(1), 551–556. https://doi.org/10.33545/26174693.2024.v8.i1h.437
Singh, S., Vij, S., & Kushwah, A. (2026). Harnessing speed breeding: a pathway to accelerating genetic gain in crop breeding. In Journal of Plant Biochemistry and Biotechnology. Springer. https://doi.org/10.1007/s13562-026-01044-4
Song, Y., Yuan, X., Wang, P., Wang, Z., Kang, L., Yang, J., Yang, G., & Hong, D. (2026). Rapid improvement of seed weight and yield by combining QTL pyramiding with speed breeding in Brassica napus L. Oil Crop Science, 11(1), 83–91. https://doi.org/10.1016/j.ocsci.2026.01.006
Terada, K., Kamal, N. M., Sakuma, S., & Ishii, T. (2026). Speed breeding for pearl millet and wild Pennisetum species with optimized pot size, CO₂ supplementation, and ABA application. Discover Applied Sciences, 8(1). https://doi.org/10.1007/s42452-025-08041-7
Wanga, M. A., Shimelis, H., Mashilo, J., & Laing, M. D. (2021). Opportunities and challenges of speed breeding: A review. In Plant Breeding (Vol. 140, Number 2, pp. 185–194). Blackwell Publishing Ltd. https://doi.org/10.1111/pbr.12909
Watson, A., Ghosh, S., Williams, M. J., Cuddy, W. S., Simmonds, J., Rey, M. D., Asyraf Md Hatta, M., Hinchliffe, A., Steed, A., Reynolds, D., Adamski, N. M., Breakspear, A., Korolev, A., Rayner, T., Dixon, L. E., Riaz, A., Martin, W., Ryan, M., Edwards, D., … Hickey, L. T. (2018). Speed breeding is a powerful tool to accelerate crop research and breeding. Nature Plants, 4(1), 23–29. https://doi.org/10.1038/s41477-017-0083-8
Williams, K., Subramani, M., Lofton, L. W., Penney, M., Todd, A., & Ozbay, G. (2024). Tools and Techniques to Accelerate Crop Breeding. In Plants (Vol. 13, Number 11). Multidisciplinary Digital Publishing Institute (MDPI). https://doi.org/10.3390/plants13111520
Xu, Y., Luo, H., Zhang, H., Yung, W. S., Li, M. W., Lam, H. M., & Huang, C. (2023). Feeding the world using speed breeding technology. In Trends in Plant Science (Vol. 28, Number 3, pp. 372–373). Elsevier Ltd. https://doi.org/10.1016/j.tplants.2022.12.003
Zainuddin, F., Ismail, M. R., Hatta, M. A. M., & Ramlee, S. I. (2024). Advancement in modern breeding and genomic approaches to accelerate rice improvement: speed breeding focus. In Euphytica (Vol. 220, Number 7). Springer Science and Business Media B.V. https://doi.org/10.1007/s10681-024-03353-y
Related posts:
 Genotyping-by-Sequencing (GBS): A Strategic Framework for High-Throughput Plant Breeding and Genomics
Genotyping-by-Sequencing (GBS): A Strategic Framework for High-Throughput Plant Breeding and Genomics
 Panama Wilt: Understanding and Managing the Banana Killer
Panama Wilt: Understanding and Managing the Banana Killer
 Bacterial Cellulose: A Game-Changer for Plant Wound Healing and Regeneration
Bacterial Cellulose: A Game-Changer for Plant Wound Healing and Regeneration
 Gene Gun Gets a Major Upgrade: Flow Guiding Barrel Boosts Plant Genome Editing Efficiency
Gene Gun Gets a Major Upgrade: Flow Guiding Barrel Boosts Plant Genome Editing Efficiency
 Unlocking Nature’s Toolkit: How Plants Use Hormones to Build Organs for Symbiosis and Survival
Unlocking Nature’s Toolkit: How Plants Use Hormones to Build Organs for Symbiosis and Survival
